
洛氏鱥染色体核型分析
2016-11-11 10:38姜玉声王茂林
广东海洋大学学报 2016年3期
张 帅,姜玉声,王 丰,王茂林 ,周 贺
(大连海洋大学农业部北方海水增养殖重点实验室,辽宁 大连 116023)
洛氏鱥染色体核型分析
张 帅,姜玉声,王 丰,王茂林 ,周 贺
(大连海洋大学农业部北方海水增养殖重点实验室,辽宁 大连 116023)
以洛氏鱥(Phoxinus lagowskii)肾组织为材料,采用腹腔注射植物血球凝集素(PHA)、秋水仙素和空气干燥法制备染色体标本,进行染色体组型分析。结果表明:洛氏鱥染色体为50条,核型公式为2n = 50 = 24m + 14sm + 12t,臂数 NF = 88 ,未发现异型染色体、随体染色体和次缢痕。
洛氏鱥;染色体;核型
洛氏鱥(Phoxinus lagowskii)隶属于鲤形目Cypriniformes鲤科Cyprinidae雅罗亚科Leuciscinae鱥属Phoxinus,曾用名长尾鱥,俗称柳根鱼,为江河野生的小型经济鱼类,分布于黑龙江、图们江、辽河、鸭绿江和黄河中上游的支流水域[1]。洛氏鱥体较长,稍侧扁,纺锤形,背部灰黄色,自背部正中至尾柄有不明显的黑条纹,腹部和体侧银黄色或银白色[2]。该鱼肌肉鲜美,肉质细嫩,肌间刺较少,蛋白质和不饱和脂肪酸含量高,市场价格较高,且具有生长速度快、适应范围广、抗病力强、对环境和温度的适应性强等特点,适于高密度大规模的池塘养殖,有广阔的发展前景[1]。
染色体作为遗传信息的载体具有种的特异性,不同物种有其特定的染色体组型,染色体组型在很大程度上反映了物种的进化历史和种间的亲缘关系。因此染色体组型研究成为细胞遗传学不可缺少的部分,对种质资源研究有极其重要的意义[3]。
目前,关于洛氏鱥的报道多局限于繁殖生物学和肌肉营养成分的研究[1,4,5]。王茂林等[1]研究了洛氏鱥个体繁殖力,康鑫等[4]分析了洛氏鱥幼鱼栖息地适宜度指数,张永泉[5]对雌雄洛氏鱥的肌肉营养成分进行比较分析。但洛氏鱥细胞生物学研究并不多见,关于其染色体核型赵守城[6]在1982年有过报道,实验材料取自河北省太原虹琳鱼场的流水渠,其水域与东北水域存在地理隔离,染色体组型可能有差异。本研究分析东北水域的洛氏鱥染色体组型,以期了解洛氏鱥遗传背景,为开展洛氏鱥的人工育苗提供细胞遗传学参数,也为其种质资源开发及演化地位研究提供理论依据。
1 材料与方法
1.1 材料
实验用鱼,9尾,体质量25~45 g,体长100 ~130mm,于2015年5月采自辽宁省本溪市太子河,暂养于大连海洋大学农业部北方海水增养殖重点实验室容积50 L的水族箱内,实验用水为曝晒的自来水,水温 26~28℃,pH 6.5~7.5,硬度80~98 mg/L。实验鱼体侧自头后缘至尾鳍基具一黑色条带(图1),尾柄长/尾柄高大于2,小于3。将该鱼线粒体控制区DNA序列与NCBI上的洛氏鱥同源序列比对,相似度为99%。因此,可断定实验鱼为洛氏鱥。
1.2 实验方法
染色体标本制备参照李雅娟等[7]方法,稍作改进。每尾鱼以鱼体质量10 μg/g的剂量腹腔注射植物血球凝集素(PHA),20 h后再次注射,剂量相同。4 h后,以8 μg/g的剂量注射秋水仙素,2.5 h后,断尾取血。剖取肾脏、鳃,用7 g/L的生理盐水冲洗2次,放入0.075 mol/L 的 KCl低渗溶液中处理60 min,用新配制的卡诺固定液固定3次,每次15~20 min。用新配制卡诺固定液于- 20℃条件下保存12 h以上。鱼组织加入 1~2 滴体积分数为50%的冰乙酸,剪碎,加50~100 μL卡诺固定液,过孔径50 μm的滤网,吸取滤液,以50~60 cm的高度滴于预冷载玻片上,自然干燥。用体积分数为10%的Giemsa染液染色 2 h,用纯水冲洗,干燥后镜检。
1.3 染色体核型分析
取雌、雄鱼各10尾共计105个分散良好的分裂相计数,以确定染色体众数。选取长度适中、着丝粒清晰、染色体众数与上述统计结果相一致的中期分裂相拍照,用 Excel 软件进行统计分析。
染色体分类采用 Levan 等[8]的标准,即按臂比将染色体分为4组:臂比为1.0~1.7的为中部着丝点染色体(m);臂比为1.8~3.0为亚中部着丝点染色体(sm);臂比为3.1~7.0的为亚端部着丝点染色体(st);臂比为7.1~8.0的为端部着丝点染色体(t)。
2 结 果
2.1 染色体数目
雌雄105个洛氏鱥肾细胞中期分裂相结果如表1。表1可见,染色体数目为50的出现频率最高为82.8%,占绝对优势,可判断洛氏鱥二倍体染色体数为50。

表1 洛氏鱥染色体数目的分布频率Table 1 Number frequency of chromosomes in Phoxinus lagowskii
2.2 染色体核型
图2表明,雌、雄洛氏鱥染色体数目和形态无明显差异。通过测量分析多个分裂良好、着丝点清晰的分裂相,获得洛氏鱥染色体的相对长度、臂比和染色体类型(表2)。按Levan等[8]的标准,洛氏鱥50条染色体中,24条为亚中部着丝粒染色体(sm),14条为亚端部着丝粒染色体(st),12条为端部着丝粒染色体(t),核型公式为2n = 50 = 24m + 14sm + 12t,臂数NF = 88。未发现异型的性染色体、随体染色体和次缢痕。但发现一对大型端部着丝粒染色体(t),相对长度为(6.00 ±0.24)%,远大于其他染色体对。
3 讨 论
鱥属鱼类分布于北美、欧洲和亚洲北部。国外对鱥属鱼类的染色体已有报道,其二倍体染色体数亦为50(表3),因此2n = 50可能是鱥属鱼类的原始核型[9]。由表3可知,洛氏鱥染色体核型与北美洲的黑带鱥和红腹鱥相似度极高,极可能来自同一分支。鱥属鱼类喜温度低、水流湍急的流水,在一些山涧溪流中往往是绝对优势种群,有的山涧溪流中甚至仅有鱥属鱼类。它们因可适应此类极端环境,生存压力低,故染色体核型有保守性。而欧洲的真鱥染色体核型多样性较大,可能与欧洲复杂的水系有关。Gold等[9]研究北美洲雅罗鱼亚科9属某些鱼类核型,发现它们有一对大型的端部或近端着丝点染色体,这或可作为鱥属鱼类的核型特征之一。

图1 洛氏鱥Fig.1 Phoxinus lagowskii
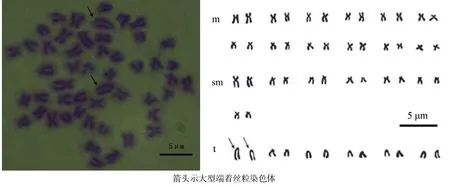
图2 洛氏鱥的染色体中期分裂相和染色体核型Fig.2 Chromosomes at metaphase and karyotypes in Phoxinus lagowskii
赵守成[6]于 1981年分析了洛氏鱥的染色体核型,其实验鱼取自河北省太原虹琳鱼场的流水渠,核型公式为2n = 48 = 24m + 4st + 10t,NF = 72,与本研究的核型有差异,可能是由染色体多态性造成的。《中国淡水鱼类染色体》[15]曾对中国淡水鱼类的染色体核型进行初步统计,仅在染色体核型上存在差异的种类就有47种,占所统计鱼类的27.16%。可见,鱼类染色体的多态现象较为普遍。Chen等[16]提出,染色体通过罗伯逊易位、臂间倒位、缺失等染色体畸变可获得新核型,具有新核型的个体在不同地理环境中通过自然选择或经过遗传漂变在某一地区形成一稳定群体。当然两者染色体核型上的差异性也不排除是实验方法和操作上的原因。
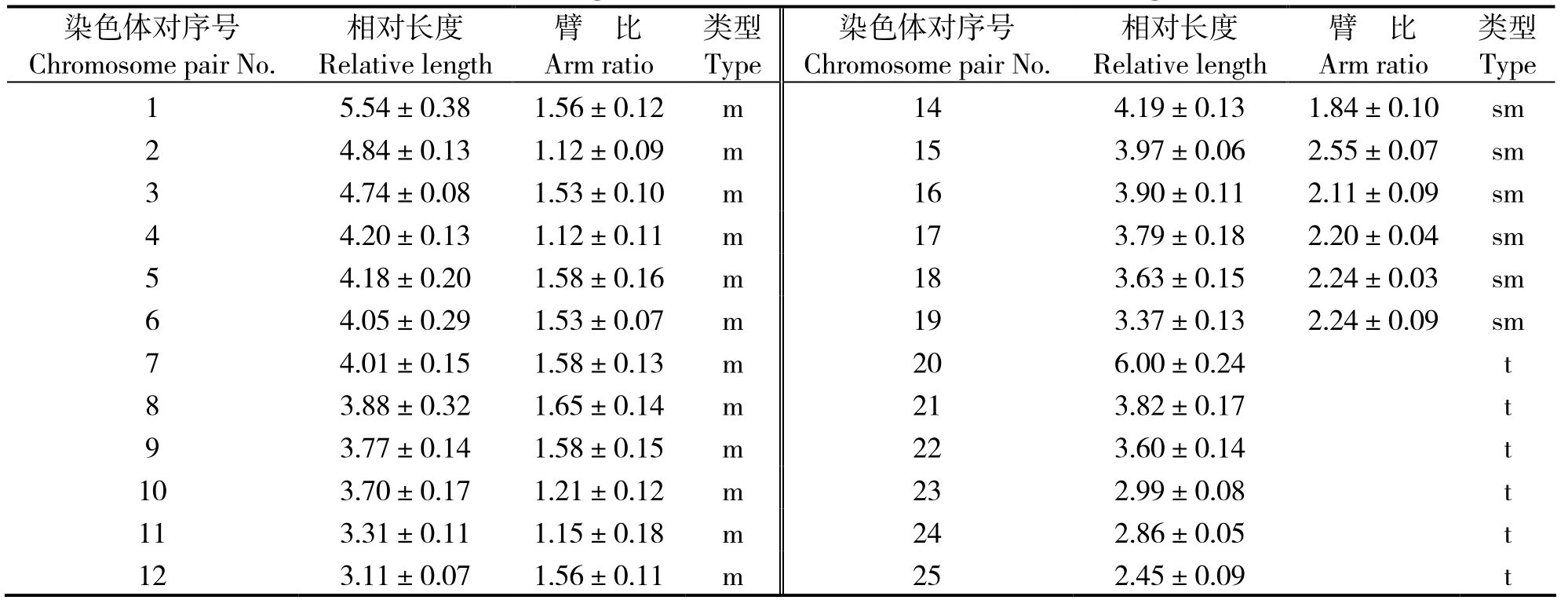
表2 洛氏鱥染色体相对长度和臂比Table 2 Relative length and ratio of chromosomes in Phoxinus lagowskii

表3 洛氏鱥与鱥属鱼类的染色体核型Table 3 Karyotype data of the genus Phoxinus
与雅罗亚科鱼类比较,洛氏鱥染色体核型与雅罗鱼属3种雅罗鱼染色体核型的相似度最高,二倍体染色体数均为50,仅染色体臂数有差异(表4)。小岛吉雄[17]对 70余种鱼类染色体数目的统计分析表明,进化上愈处于上位的类群,染色体数目的离散度愈小。即物种越处于进化上位,染色体越收敛,端部着丝粒染色体越多,臂数(NF)越少,在系统进化上属高位类;反之,则属低位类。可见,在系统进化上,洛氏鱥相对于圆腹雅罗鱼(Leuciscus idus)处于上位类,相对于瓦氏雅罗鱼(L.waleckii)和准噶尔雅罗鱼(L.merzbaeher)则处于下位类。鯮鱼(Bambusa)和鱤鱼(Bambusa)的染色体二倍体个数为48,可能是由2n = 50经罗伯逊易位而形成。马口鱼(Bidens)和宽鳍鱲(Platypus)二倍体染色体核型分别为 76、78,与洛氏鱥相差最大,可能是由2n = 48或2n = 50通过着丝点断裂方式使染色体数目大大增加。
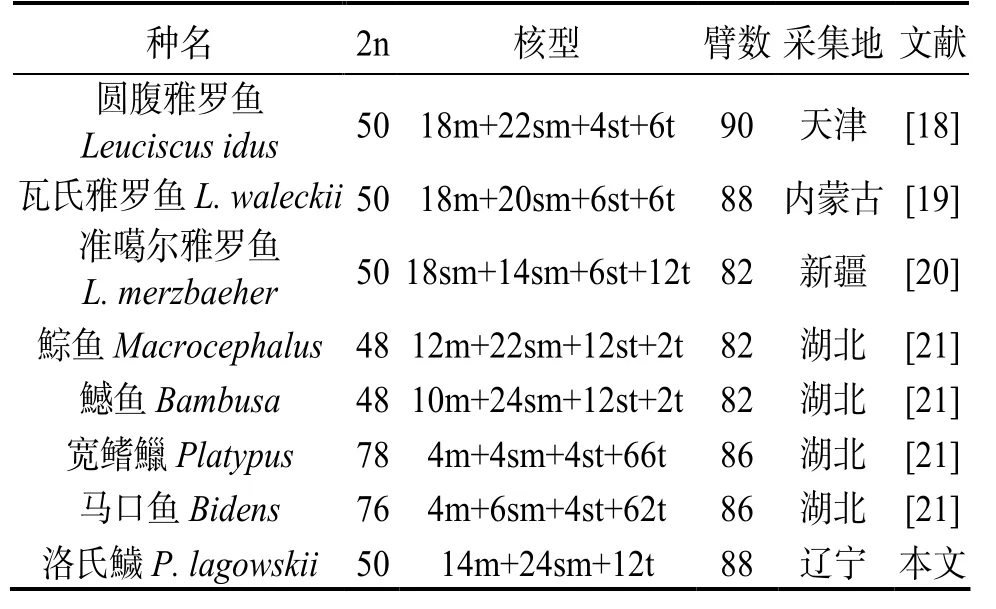
表4 洛氏鱥与雅罗亚科鱼类的染色体核型Table 4 Karyotype data of the Leuciscinae
鱼类在脊椎动物中属原始类群,性染色体分化不明显[22]。据 Vasiliev[23]统计,在已有染色体研究的2 000多种鱼类中,具有异型性染色体的物种仅占 2.5%。本研究分别制备雌、雄洛氏鱥染色体滴片,结果表明,雌、雄洛氏鱥的染色体核型无区别,未发现异型的性染色体。
[1]王茂林,李保民,姜玉声.本溪太子河流域洛氏鱥个体繁殖力研究[J].长江大学学报(自然科学版),2013,10(10):28-32.
[2] 张觉民.黑龙江鱼类志[M].哈尔滨:黑龙江科学出版社,1995:50-52.
[3] 覃映雪,苏永全,王胜强,等.花尾胡椒鳃染色体核型分析[J].海洋学报,2003,25(4):147-150.
[4] 康鑫,张远,张楠,等.太子河洛氏鱥幼鱼栖息地适宜度评估[J].生态毒理学报,2011(3):310-320.
[5] 张永泉,尹家胜,杜佳.雌雄洛氏鱥肌肉营养成分的比较分析[J].食品科学,2013,34(17):259-262.
[6] 赵守城.鱼岁鱼染色体组型分析[J].淡水渔业,1982,25(4):147-150.
[7] 李雅娟,钱聪,印杰,等.不同倍性泥鳅杂交后代染色体数目组成的研究[J].大连海洋大学学报,2012,27(4):326-332.
[8] LEVAN A,FREDGA K,SANDBERG A.Nomenclature for centromeric position on chromosomes[J].Hereditas,1964,52:201-220.
[9] GOLD J R,WOMAC W D,DEAL F H.Cytogenetic studies in North American minnows(Cyprinidae).ⅦKaryotypes of thirteen species from the southern United States[J].Cytologia,1981,46(1/2):105-115.
[10] BERBEROVIC L,SOFRADZIJA A.Basic data about diploid chromosome complement of phoxinuns(Cyprinidae:Pisces) [J].Bull Scient Cons Acads PSF Yugosl,(A),1974,19(5/6):142-143.
[11] STEFANO Cataudella,LUCIANA Sola,ROSANNA Accame Muratori.The chromosomes of 11 species of Cyprinidae and one Cobitidae from Italy,with some remarks on the problem of polyploidy in the Cypriniformes[J].Genetica,1977,47(3):161-171.
[12] BOROŃ A.Comparative chromosomal studies on two minnow fish,Phoxinus phoxinus(Linnaeus,1758) and Eupallasella perenurus(Pallas,1814):an associated cytogenetic-taxonomic considerations[J].Genetica,2001,111(3):387-395.
[13] HAFEZ R,LABAT R,QUILLIER R.Some species of Cyprinidae from the midi-pyrenees region[J].Bulletin de la Societe D'histoire Naturelle de Toulouse,1978,114(1/2):122-159.
[14] JOSWIAK G R,MOORE W S.Karyotypes of three species of the genus Phoxinus (Pisces:Cyprinidae)[J].Copeia,1980(4):913-916.
[15]余先觉,周暾,李 康,等.中国淡水鱼类染色体[M].北京:科学出版社,1989:149-166.
[16]CHEN T R,EBELINGET A W.Chromosomes of the goby fishes in the genus Gillichthys[J].Copeia,1971,1971(1):171-174.
[17] 小岛吉雄.鱼类细胞遗传学[M].广州:广东科技出版社,1990:12.
[18] 金万昆,赵宜双,张慈军,等.圆腹雅罗鱼的染色体核型分析[J].内陆水产,2009(1):56-61.
[19] 张锦霞,王祖熊.瓦氏雅罗鱼染色体组型研究[J].水生生物学报,1987,11(4):367-370.
[20] 王佳君,胡文革,孔磊.准噶尔雅罗鱼染色体核型及带型的初步研究[J].动物学杂志,2010,45(6):120-126.
[21] 李渝成,李康,洪云汉.中国鲤科鱼类染色体组型的研究:雅罗鱼亚科 7 种鱼的染色体组型分析及有关鲤科鱼类系统关系的探讨[J].遗传学报,1985,12(5):367-372.
[22] 童金苟,朱嘉濠,关海山.鱼类性别决定的遗传基础研究概况[J].水产学报,2003,27(2):169-176.
[23] VASILIEV V B.Evolutionary karyology of fishes [M] // Institute of evolutionary morphology and ecology.USSR Academy of Sciences(in Russian).Moscow:Publ Nauka,1985:16.
(责任编辑:刘庆颖)
Karyotype of Phoxinus lagowskii
ZHANG Shuai,JIANG Yu-sheng,WANG Feng,WANG Mao-lin,ZHOU He
(Key Laboratory of Mariculture & Stock Enhancement in North China's Sea,Ministry of Agriculture,Dalian Ocean University,Dalian 116023,China)
Karyotype of Phoxinus lagowskii was studied with chromosome samples prepared from kidney by injecting phytohaemagg lutinin(PHA) and colchicine and air-dried method.The results showed that there were 50 chromosomes in P.lagowskii,and karyotype formula of P.lagowskii was 2n = 50 = 24m + 14sm + 12t; NF = 88.There was no evidence of heterotypical chromosome,satellites or secondary constriction.
Phoxinus lagowskii; chromosome; karyotype
Q959.46+8.3
A
1673-9159(2016)03-0106-04
10.3969/j.issn.1673-9159.2016.03.017
2015-11-11
辽宁省“三区”人才计划项目(500215201);大连海洋大学校列项目(100914198)
张帅(1988-),男,硕士,从事水产集约化养殖研究。E mail Zhangshuai1900@126.com
王茂林(1980-),男,副教授,从事水产养殖学、鱼类生理生态研究。E mail wml13804@126.com
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
生物技术进展(2022年1期)2022-02-11
宁夏医学杂志(2020年3期)2021-01-21
儿童时代·幸福宝宝(2020年9期)2020-09-08
科学之谜(2018年9期)2018-12-17
中国当代医药(2015年9期)2015-03-01
郑州大学学报(医学版)(2015年1期)2015-02-27
现代检验医学杂志(2015年4期)2015-02-06
遗传(2015年4期)2015-02-04
台湾农业探索(2012年6期)2012-02-15
