
红花双列杂交后代羟基黄色素A含量的遗传效应分析
2020-12-31 05:58梁慧珍许兰杰余永亮谭政委杨红旗董李春明刘新梅张收良
作物杂志 2020年6期
梁慧珍 许兰杰 余永亮 谭政委 杨红旗董 薇 李 磊 李春明 刘新梅 张收良
(1河南省农业科学院芝麻研究中心,450002,河南郑州;2河南省农业科学院西峡分院,474550,河南西峡)
红花(Carthamus tinctoriusL.)是菊科管状花亚科红花属植物中唯一的栽培植物[1],自汉代起,就有其栽培和药用记载[2]。红花既是药用植物,又是油用和工业用植物,全世界每年种植红花约110万hm2。中国是红花种植大国,年种植面积3万~4万hm2,主要分布在新疆、云南和河南等地[3]。红花的主要药用成分红花黄色素(saffower yellower)和红花苷(crocin)具有抗炎、镇痛和抗肿瘤等作用。含有红花成分的药物具有增强冠状动脉血流量、抗心肌缺血,降血脂和抗血栓的功能[4-7]。红花品质育种的主要目标是提高羟基红花黄色素A(hydroxy safflor yellow A,HSYA)含量。目前红花的新品种选育方法以系统选育和杂交育种为主。杂交育种的关键技术是选择杂交亲本和杂交后代群体。杂交材料的遗传力、配合力效应已广泛应用于水稻[8]、玉米[9]、大豆[10]、蚕豆[11]和红松[12]等植物数量性状的遗传和育种研究,但应用到HSYA含量的研究尚未见报道。吴沂芸等[13]对红花黄酮类成分累积量与功能基因表达水平进行了关联分析,证实了关键酶的催化作用,可以通过调节chs与chi的表达来提升红花中HSYA含量;许兰杰等[14-15]研究了HSYA的动态积累规律和热稳定性,及其与花色的相关性,认为温度和时间对HSYA含量均存在较大影响,并且发现HSYA与红花的颜色有关,呈现以下规律:红色红花的HSYA含量高于橙色、黄色和白色红花;Liu等[16]利用分子标记方法对新疆和云南红花品种进行DNA分析,筛选出了12对相关序列扩增多态性(sequence-related amplified polymorphism,SRAP)引物组合;唐洁[17]建立了红花的纯化mRNA模板,反转录合成cDNA反应体系,筛选出2组多态性引物,但基于胚、细胞质和母体遗传效应3套遗传体系研究HSYA的遗传机理尚未见报道。本研究配制亲本组合时选用了6个河南当地HSYA含量差异较大的红花品种,运用双子叶植物种子数量性状的遗传模型和统计分析方法[18-20],利用SPSS 19.0软件分析控制HSYA含量三套不同遗传体系下的胚、细胞质和母体植株的基因效应和环境互作效应,预测其遗传效应值,评价杂种后代的杂种优势和亲本的遗传力,摸索HSYA含量的遗传规律,为红花品质数量性状的种质改良与优异性状鉴定提供理论依据。
1 材料和方法
1.1 试验材料
供试红花种子分别是豫红花1号(P1)、延津红花A(P2)、延津红花B(P3)、原阳红花A(P4)、延津红花C(P5)和原阳红花B(P6)。P1、P2和P33个亲本为较高HSYA含量品种,P4、P5和P6亲本为中低HSYA含量品种。所有参试样本均为二倍体红花。来源于河南省农业科学院红花资源库,材料种植于河南省农业科学院现代农业科技试验示范基地(以下简称“原阳示范基地”)。
1.2 试验设计
根据双子叶植物数量性状的遗传模型和统计分析方法[18-20],在2014年用上述亲本在原阳示范基地选取了亲本及只进行正交组合的完全双列杂交方法配制的杂交组合,于2015年获得F1代杂交种子;2015年和2016年分别种植亲本和F1,每年都按照同样的亲本配制杂交组合,得到杂交种子F1和F2。该试验采取随机区组排列,单行区,3次重复,行长5m,行距0.4m,株距0.14m。
1.3 HSYA含量测定
1.3.1 采样 对红花花丝进行HSYA含量测定。采样时间为红花开花后第3天,采摘时间为8∶00-9∶00。将样品在50℃烘箱内烘干,烘干时间为240min[14]。
1.3.2 色谱条件 参考许兰杰等[14]的方法:设置色谱柱Agilent C18柱(25.0mm×4.6mm,5μm);流动相中乙腈(C2H3N)、甲醇(CH3OH)和浓度0.7%的磷酸溶液(H3PO4)的比例为2∶26∶72;检测波长403nm;柱温30℃;流速1.0mL/min;进样量9μL。
1.3.3 对照溶液和样品溶液的制备 参考赵明波等[21]的方法并略作改变。对照溶液:称取HSYA对照品10.0mg,放入10mL容量瓶中,加入甲醇溶液(浓度25%)溶解、定容,成为1.0mg/mL的对照溶液,用0.45μm微孔滤膜过滤后备用。样品溶液:称取红花粉末样品各100mg,置于具塞锥形瓶中,加入浓度25%甲醇溶液10mL称重,25℃超声提取1h,放冷再称重,用25%甲醇溶液补足减失的质量,摇匀,过滤,再用0.45μm微孔滤膜过滤后备用。
1.3.4 HSYA含量的测定 分别吸取对照溶液和供试样品溶液20μL注入液相色谱仪,按2005版《中国药典》(一部)规定的方法测定HSYA的峰面积,采用外标法计算HSYA含量。
1.3.5 HSYA标准曲线绘制 分别吸取对照溶液5、10、15、20、25、30和35μL,测定HSYA的峰面积,绘制标准曲线。
1.4 精密度试验
取同一生药样品,按照样品溶液的制备方法制备,连续进样5次,测定HSYA的峰面积,结果峰面积的相对标准偏差(RSD)为2.29%,表明仪器精密度良好。
1.5 重现性试验
取同一批样品6份,按样品溶液制备方法制备,测定HSYA含量,结果其RSD为2.37%,表明该方法重现性良好。
1.6 数据分析
利用Excel 2019对亲本和F1、F2代HSYA含量进行统计分析;采用双子叶植物遗传模型和统计分析方法[18-20]中胚、细胞质和母体植株3套遗传体系分析HSYA含量的基因主效应、基因型与环境互作效应。利用世代平均数估算遗传主效应方差VG、基因型×环境互作方差VGE和机误方差Ve。HSYA的表现型方差VP=VG+VGE+Ve。其中:VG=VA+VD+VC+VAm+VDm,VGE=VAE+VDE+VCE+VAmE+VDmE。HSYA 的 遗传率h2=hG2+hGE2。其中,hG2=hGo2+hGc2+hGm2,hGE2=。VA、VD和VC分别代表胚加性方差、胚显性方差和细胞质方差,VAm和VDm分别代表母体加性方差和母体显性方差;VAE、VDE和VCE分别代表胚加性效应、胚显性效应和细胞质效应3种效应下的环境互作方差,VAmE和VDmE分别代表母体加性效应和母体显性效应的环境互作方差。hG2为普通遗传率,hGo2、hGc2和hGm2分别代表胚普通遗传率、细胞质普通遗传率和母体普通遗传率;hGE2为互作普通遗传率,hGoE2、hGmE2和hGcE2分别为胚互作遗传率、母体互作遗传率和细胞质互作遗传率。
采用调整无偏预测法[18-20]预测胚加性效应(A)、胚显性效应(D)、细胞质效应(C)、母体加性效应(Am)、母体显性效应(Dm)和相应的环境互作效应(AE,DE,CE,AmE和DmE)。分析预测值的大小和方向,评判亲本的育种价值,预测组合的杂种优势。HSYA的杂种优势总量H=HG+HGE。各杂种优势分量间的关系为:HG=HGo+HC+HGm,HGE=HGoE+HCE+HGmE。其中,HG为普通优势,HGo、HC和HGm分别代表胚普通优势、细胞质普通优势和母体普通优势;HGE为环境互作优势,HGoE、HCE和HGmE分别代表胚互作优势、细胞质互作优势和母体互作优势。
2 结果与分析
2.1 亲本及杂种后代HSYA含量的平均表现
由表1可知,亲本间HSYA含量差异较大,亲本P1、P2和P3含量较高,亲本P4、P5和P6含量较低。杂种F1和F2间HSYA含量差异也比较大。F1代HSYA含量的变幅为10.03~26.19mg/g,F2代为7.64~29.36mg/g。分析F1和F2群体值发现,F1P1、F1P4和F1P5子代材料HSYA平均含量均高于亲本,表现出正向的杂种优势。F1P2和F1P3子代材料HSYA平均含量均低于亲本,表现出负向的杂种优势。F2代材料HSYA平均含量均低于F1代,主要是由于F1表现出杂种优势后,F2代出现分离引起的。
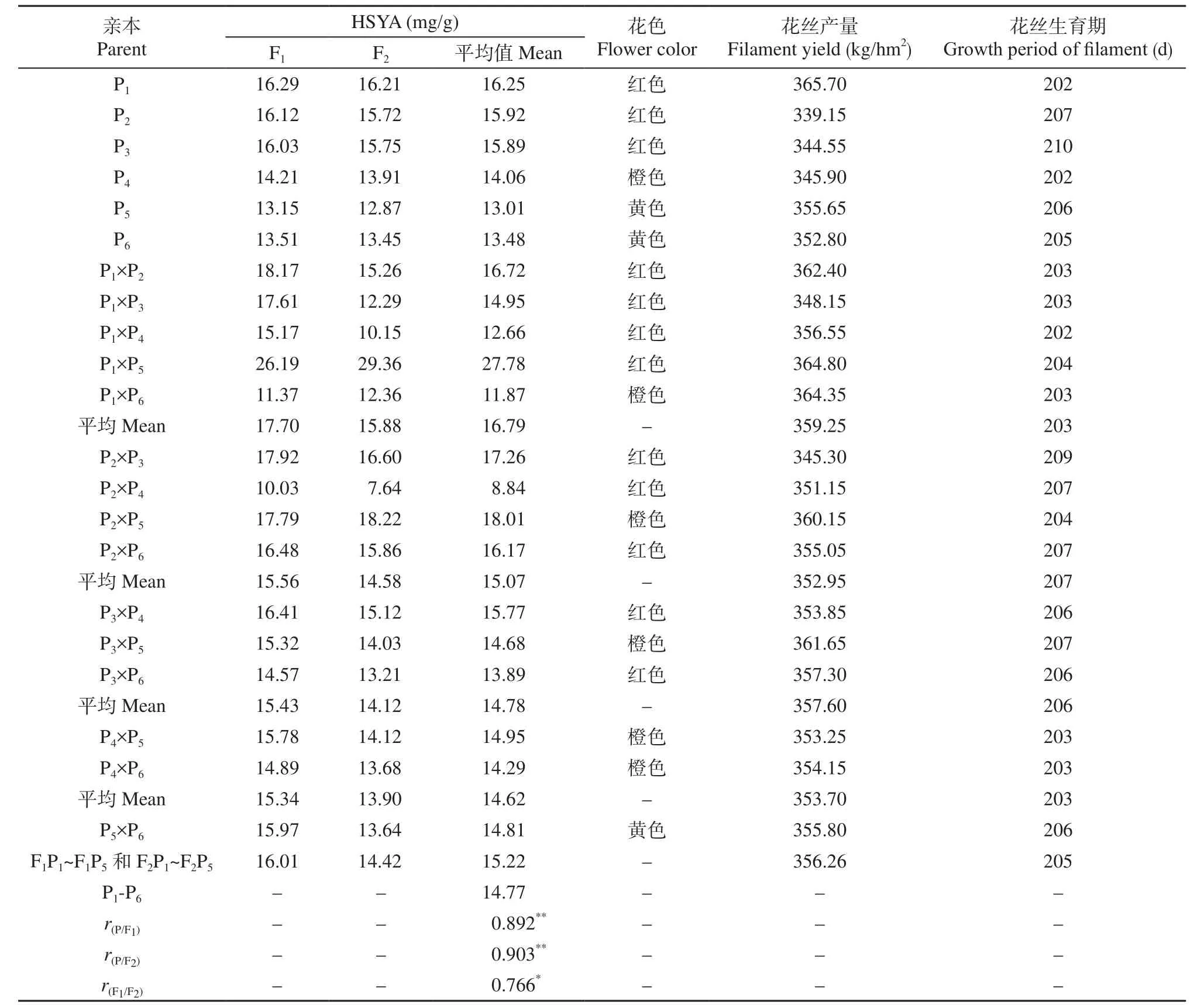
表1 亲本和F1、F2代HSYA含量和农艺性状的平均值Table 1 Means of HSYA content and agronomic traits of parents and relative F1, F2 in Carthamus tinctorius
对比亲本与后代发现,F1和F2的HSYA含量高低与亲本含量的高低相对应。相关系数分析表明,亲本与F1、F2之间、F1和F2之间均表现出极显著或显著正相关。P1、P2和P3亲本中HSYA含量高,其后代材料中HSYA含量相对较高。说明HSYA含量可能受胚加性遗传、母体或细胞质遗传体系的控制。同时发现,P5亲本HSYA含量较低,作亲本的后代材料中HSYA含量相对较高,表现出超亲现象,说明P5亲本中HSYA含量受加性效应遗传控制的同时,还受到环境互作因素的影响,表现出较强的杂种优势。结果显示,P1×P5杂交后代中HSYA的含量最高。所以,在改良和选育高HSYA红花品种时,优先选择HSYA含量高的亲本作母本,选择杂种优势大的作亲本。
2.2 HSYA含量的遗传效应与方差分量估算
分析表2中各个方差分量发现,遗传主效应方差(VG=VA+VD+VC+VAm+VDm=0.127)和环境互作方差(VGE=VAE+VDE+VCE+VAmE+VDmE=0.044)分别占遗传方差总量(V=VG+VGE=0.171)的74.27%和25.73%,说明HSYA含量同时受遗传主效应和基因型与环境互作效应的影响,但以遗传主效应控制为主。同时发现,多数遗传方差分量达极显著水平,说明HSYA含量的遗传受上述3套不同遗传体系基因效应的影响。而且HSYA含量的遗传以母体遗传效应(VAm+VDm+VAmE+VDmE)为主,占遗传方差总量VG的60.82%;胚效应(VA+VD+VAE+VDE),占遗传方差总量VG的29.24%;细胞质效应(VC+VCE)仅占VG的9.94%。说明基因的加性效应和胚效应是控制HSYA含量的主要遗传效应。在各遗传方差分量中,母体加性方差VAm=0.049**,数值最大,说明杂种后代HSYA含量在单株上变异不大,通过母体植株的总体表现可以选择出高HSYA含量的种质材料。母体加性互作方差VAmE为0.019**,数值较大,说明配制杂交组合时,优先考虑双亲的选择,特别是母本的选择更为重要,环境对母体植株基因的遗传也有一定的影响。正确选择基因的加性效应,遗传选择过程中可以形成累加效应,在母本中可以传递细胞质效应,两种效应直接影响后代的选择结果。胚显性方差VD=0.042**,胚显性互作方差VDE=0;母体显性方差VDm=0.036**,母体显性互作方差VDmE=0,说明杂种后代中同时产生了种子杂种优势和母体杂种优势,且两种杂种优势的主效应基因均不受环境影响。机误方差Ve=0.192**,达极显著水平,说明杂交后代中HSYA含量还受到环境机误或抽样误差的影响。
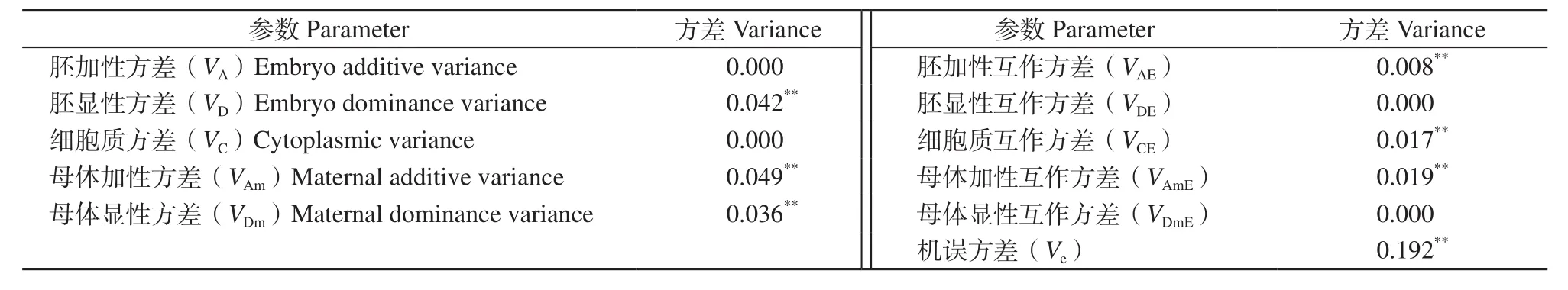
表2 HSYA含量的遗传方差分量估计值Table 2 Estimation of genetic variance components for HSYA content in Carthamus tinctorius
2.3 亲本遗传效应值的预测
杂交亲本的育种价值决定了品种HSYA含量的改良和选育效果(表3)。表3中几乎所有亲本的胚加性效应、母体加性效应和细胞质效应预测值都达到显著水平,遗传效应值为正,说明3套遗传体系的基因效应对杂种后代中HSYA含量增加起到正向作用,反之,则起到负向作用。P1亲本GⅠ和GⅡ分别为0.745和0.285,表现最佳。预计以P1作为亲本,对后代中HSYA含量的改良效果比较理想。P2、P3和P6在2年中的遗传总效应分别为–0.136、–0.222、–0.203(2015 年)和 –0.038、–0.159、–0.176(2016年),对杂种后代的HSYA含量起负向作用。P4和P5在2年中的遗传总效应表现不同,一个年份为正值,另一个年份为负值。
2.4 杂交组合显性效应预测和杂种优势分析
杂交育种是提高作物品质的重要途径。除胚加性效应和细胞质效应的作用外,基因显性效应也对作物的杂种优势发挥着重要作用,可以通过对F2显性效应分析判断其杂种优势表现。
表4表明,21个红花组合的胚显性效应平均值为0.0016,变异范围为–0.6325~0.3752,其中,14个组合的胚显性效应值为正(0.0052~0.3752),杂种优势对F2中HSYA含量增加起正向作用;7个组合的胚显性效应值为负(–0.7765~–0.0612),杂种优势对F2代中HSYA含量增加起负向作用。2年的显性互作效应平均值分别为0.0382和0.1006,对杂种F2代HSYA含量增加起正向作用。在21对杂交组合中,P3×P4胚显性效应值(0.3752)最大,2年胚显性互作效应分别为0.0413和0.0106,说明胚显性效应和胚显性互作效应可以增加后代HSYA含量。同时,P1×P5组合表现出的杂种优势与此类似,仅数值略偏低。
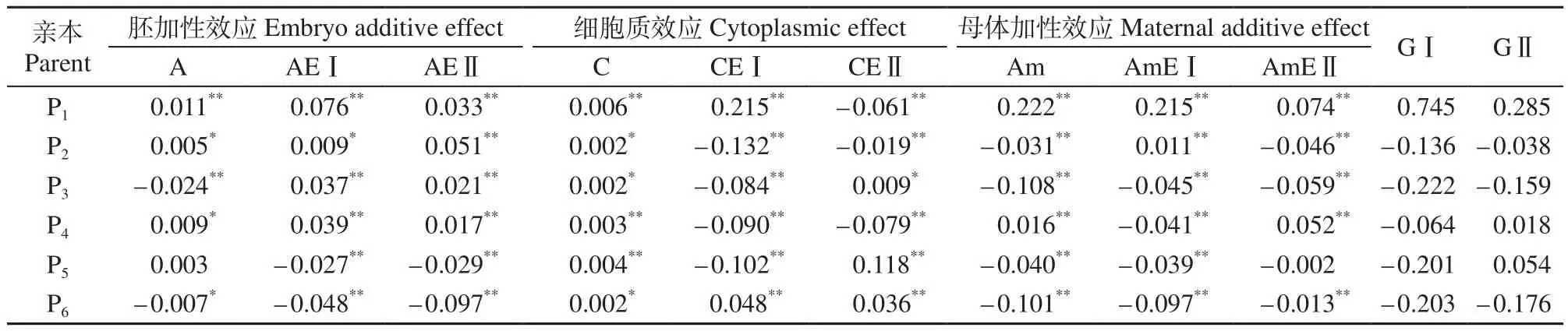
表3 HSYA含量的遗传效应预测值Table 3 Predicted genetic effects of HSYA content in Carthamus tinctorius mg/g
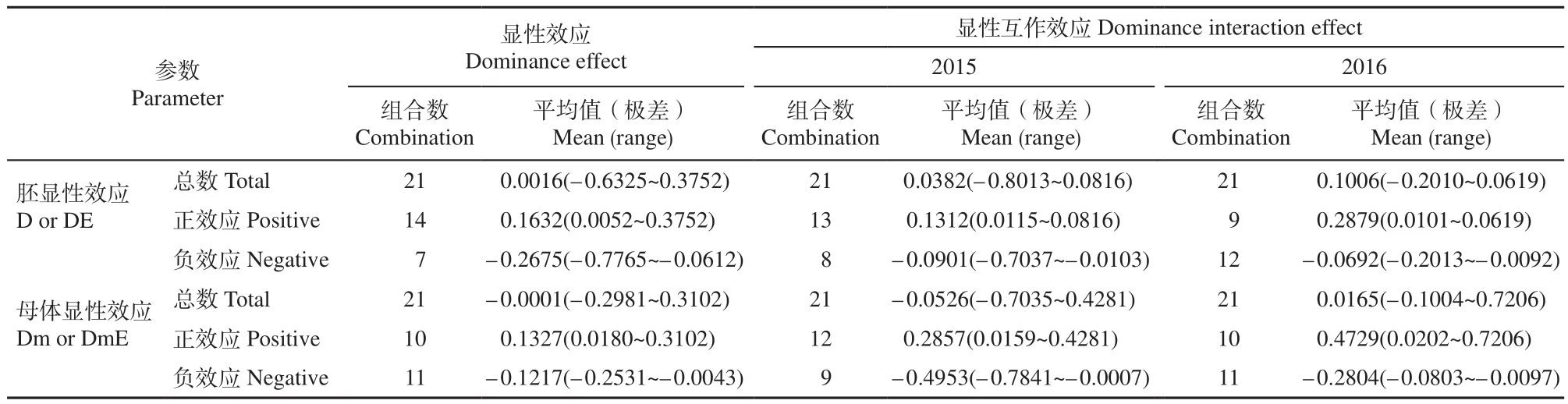
表4 F2代HSYA含量的显性效应预测值(极差)Table 4 Predicted dominance effects (range) for HSYA content of F2 generation mg/g
21个组合母体显性效应的平均值为–0.0001,变异范围为–0.2981~0.3102。11个组合母体显性效应为负值(–0.2531~–0.0043),10个组合母体显性效应为正值(0.0180~0.3102),杂种优势在不同的组合中对F2代HSYA含量的影响不同,实际工作中需要选择效应值为正值的杂交组合。2015年和2016年母体显性互作效应分别为–0.0526和0.0165,杂种优势对F2代HSYA含量的影响各有不同。因而,母体显性互作效应对HSYA含量的影响受环境影响较大。为筛选出增加F2代中HSYA含量的最佳组合,对所有参试杂交组合比较,发现P1×P5组合的母体显性效应最大(0.3102),2年母体显性互作效应均为正值,表现最为显著。
综合考虑胚显性效应、母体显性效应和显性互作效应的共同作用,P1×P5组合综合表现最佳,推荐用于高HSYA含量红花育种实践。
3 讨论
3.1 HSYA含量的遗传改良
数量性状遗传同时受多种遗传系统基因效应和基因型与环境互作的控制[22]。双子叶作物种子中胚营养物质来源于母体植株,但和母体植株不是同一个遗传世代[23-24]。合子分化发育成胚胎,最终发育成正常植株,含有全部遗传信息[9]。胚基因的表达对HSYA含量等品质性状有很大影响。本研究中,高HSYA含量亲本P1的胚加性效应值最大(0.011**),达极显著水平。以P1为亲本形成的杂种后代在所有参试样品中HSYA含量平均值最高,可以作为优选亲本材料。
经典遗传学认为,3套遗传体系中,母体植株提供胚营养物质,细胞质基因通过叶绿体影响植株的光合作用,这两种效应和基因型×环境互作效应共同影响杂种后代性状的表现,但影响程度不同[25]。根据环境互作效应的表现,可以定向选择适应不同环境的育种材料[26]。因此,分析多种遗传效应对HSYA含量的影响,有利于提高杂交后代的选择效率。本研究中,细胞质效应:P1>P5>P4,P2、P3和P6较小;细胞质效应×环境互作效应:2015年,P1>P6;2016 年,P5>P6>P3,两年中其余亲本效应较小,皆为负值。综合考虑,选用P1和P5作亲本材料,可能有利于提高杂交后代的选择效率。
本研究中,母体遗传以加性效应为主,且母体植株遗传效应大于胚效应和细胞质效应,说明同一植株上的红花杂种后代种间分离不大。实际育种工作中,通过母体植株的总体表现可以选择出表现优异的红花种质,特别在低世代选择时,选择效果更为显著。同时,为提高HSYA含量的改良效果,可以综合考虑细胞质效应对HSYA遗传的影响,采用正反交育种方法予以实现。
本研究通过对不同遗传体系中基因效应的分析,阐明了HSYA含量遗传规律,对实际育种工作具有理论指导意义,对提高品种HSYA含量的改良效率及HSYA种质资源的高效利用具有一定参考价值。
3.2 亲本在HSYA性状改良中的杂种优势利用价值
杂种优势是杂合体在1种或多种性状上优于2个亲本的现象。显性假说认为双亲有利显性基因的聚合积累和相互补充作用,增加了杂合子代F1的生长势,产生杂种优势[27]。亲本显性基因越多,杂种优势表现越强。遗传力越小和上位效应越大的性状,后代的杂种优势越明显[28]。上位效应由非等位基因间互作产生,表现出2个等位基因的表现型表达产生了遮盖作用。杂种优势的表现主要与加性×显性和显性×显性2种上位性效应有关,尤其会影响低世代数量性状的选择效果[29]。Li等[30]认为水稻杂种优势主要是由上位性产生的。关于上位性对HSYA含量杂种优势的影响,将在本研究的基础上进一步深入开展。本研究初步筛选出了改良HSYA含量的亲本材料,P1亲本HSYA含量最高,是提高后代HSYA含量的理想亲本,可以在育种中加以利用。目前已经利用P1作亲本培育出一批表现良好的新品系材料。同时发现,P5亲本HSYA含量较低(13.01),遗传效应值较小,2015年为–0.201,但P1×P5母体显性效应(0.3102)最大,胚显性效应和母体显性互作均为正值,在本研究中杂种优势表现最为突出。利用P1×P5配制的杂交组合,在实际工作中已经培育出高HSYA含量品系材料,表现优异。本研究结果可为后代材料在杂种优势利用中的亲本选择提供参考。
4 结论
在HSYA含量的遗传体系中,母体遗传效应影响最大,胚效应次之,细胞质效应影响相对较小。3套遗传体系均表现出基因主效应大于环境互作效应。豫红花1号(P1)作亲本有利于增加杂交后代HSYA含量,亲本组合P1×P5有利于提高后代杂交组合HSYA的含量。
猜你喜欢
课外生活·趣知识(2022年2期)2022-02-08
蔬菜(2021年7期)2021-11-27
辽河(2021年10期)2021-11-12
山西农经(2016年5期)2016-02-28
上海农业学报(2016年5期)2016-02-10
中国蔬菜(2015年9期)2015-12-21
应用海洋学学报(2015年3期)2015-11-22
中国当代医药(2015年20期)2015-03-01
火炸药学报(2014年1期)2014-03-20
茶叶通讯(2014年2期)2014-02-27
