
光照对锦江牛维持行为及活动量的影响
2021-05-28 04:14曹锦承李婷婷杨珍珍颜培实
畜牧与兽医 2021年6期
曹锦承,李婷婷,杨珍珍,颜培实
(南京农业大学动物科技学院,江苏 南京 210095)
动物对光环境变化的反应,在行动上具有瞬时性和持续性的不同程度时间性变化,在空间上会发生位点改变[1]。外向动物行为与动物健康和生长存在内在联系,剖析动物行为变化可以折射动物环境的舒适程度[2]。
自然光在秋冬季节缩短,利用长光周期(long-day photoperiod)即延长光照时间能够显著改善动物的生产性能。在奶牛管理中应用长光周期发现16 h光照可提高奶牛生产性能,促进奶牛的采食行为[3]。在小马驹的生产管理中运用长光周期能够增加小马驹干物质消化率,改善其生长性能,增加采食频率,减少总体采食时间[4]。邱静芸等[5]发现长光周期可提高锦江牛的日增重,增加趴卧休息率,降低活动量,认为肉牛趴卧休息率的提高可以减少其非生产能量的消耗,从而提高肉牛的生产性能。
动物行为因其动机而不同。维持行为中采食、休息和修饰等行为被视为正向的,而争斗和规癖行为被视为负向强化的结果。趴卧休息率的下降会引起动物生产性能的降低[6]。奶牛在橡胶垫、粪床等较为舒服的垫料上会增加趴卧时间[7],长光周期会提高奶牛的趴卧休息时间[8],为奶牛的生产提供有利的环境。三轴加速度传感器已常规用于奶牛活动量的测定,可判定奶牛的行为特征,在放牧牛可与全球定位系统(GPS)技术配合测定行走距离与总体活动量。本试验通过加速度传感器收集锦江牛活动量数据,聚类分析锦江牛的行为量化特征。
在邱静芸等[5]研究长光周期与自然光照相比较的基础上,本研究将长光周期与稳定短光照相比,将锦江牛分为长光周期LD组(16L∶8D)和等光周期对照SD组(12L∶12D),通过观察其行为及活动量,探讨光照对肉牛采食行为、趴卧休息行为、反刍行为和活动量的影响,进一步探讨长光周期对休息行为正向强化。
1 材料与方法
1.1 试验动物与设计
试验于2018年9月至2019年1月在江西高安市裕丰农牧有限公司进行。试验牛场为东西走向的半开放式牛舍,南北侧各12个牛栏,牛栏规格为4 m×3 m×1.5 m。在牛舍的南半侧以中轴线用遮光帘分隔东西两区,每区6组牛栏,每栏散养3头,公牛(311.17±18.04)kg和母牛(152.78±21.15)kg各18头,东西两区各有9头公牛和母牛。东区SD组在自然光照的基础上人工增加光照至12 h,12L∶12D(6:00—9:00和15:00—18:00补光);LD组,16L∶8D(6:00—9:00和15:00—21:00补光)。LED灯(13 W)安装于离地2.4 m,料槽后30 cm处,相间2 m,共计24盏。预试期5 d,正式试验期96 d。
1.2 试验日粮与日常管理
2组日粮见表1,自由采食,自由饮水,每天上午6:00、11:30及下午5:30定时清理牛粪。
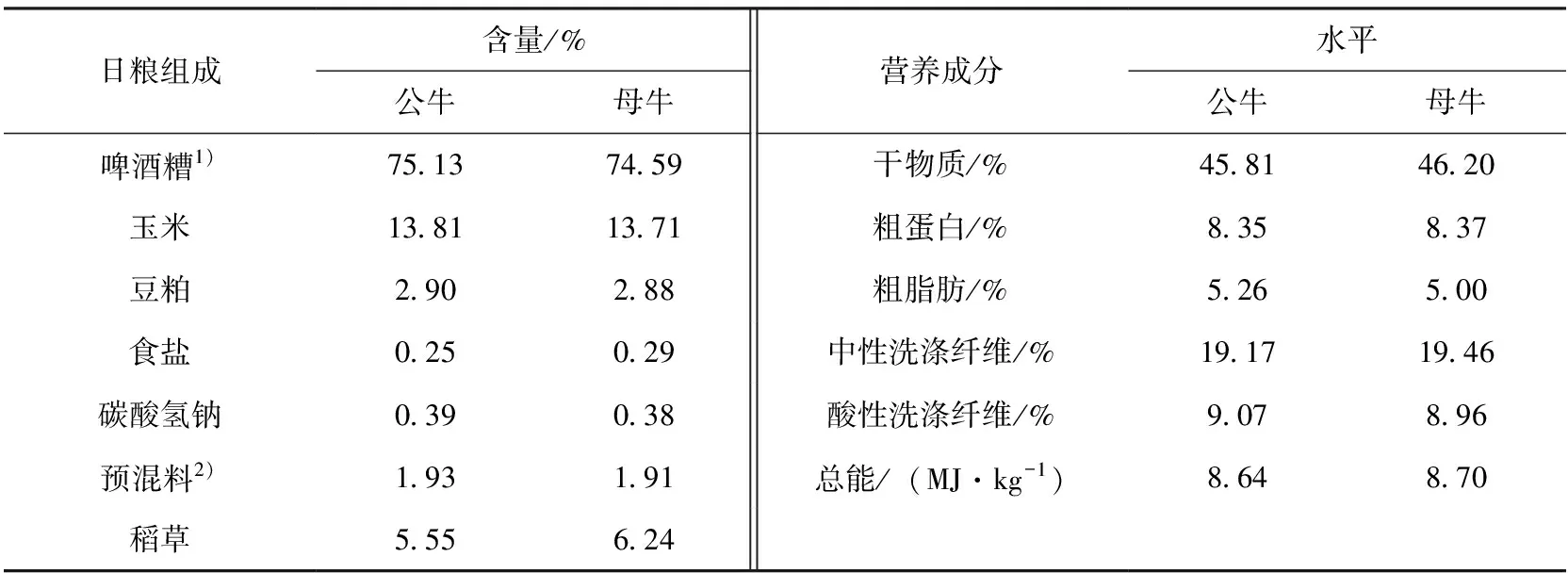
表1 试验饲粮组成及营养水平
1.3 行为观察
在试验期10月7日至10日连续4 d于不同时间点(7:00、9:00、11:00、13:00、17:00)观察并记录36头锦江牛的趴卧状态。10月19日至10月23日(光照前期)及11月27日至12月1日(光照后期),利用摄像头记录8:00—11:00和14:00—17:00锦江牛的采食行为、反刍行为、趴卧行为。
1.4 活动量的测定
在12月6日至10日上午8:00—11:00,下午2:00—5:00,每组公牛随机挑选3头,将传感器三轴加速度计量器(BWT901CL)以A型项圈的方式固定于颈侧,连续观察5 d,通过记录特征行为X、Y、Z方向的加速度数据。
K-means聚类算法可以将锦江牛的状态分为静止、微动、慢走、快走4种行为特征,表示其活动量。本试验结合项圈将三轴加速度计量器佩戴于公牛颈部,传感器的Z轴垂直指向地面,X、Y轴分别指向肉牛的前方及右侧。将4种行为特征取K等于4。试验所得数据S={X1,X2,X3,…,Xn},其中n为数据样本的个数,Xi=[xt,yt,zt],xt,yt,zt分别为X、Y、Z方向在t时刻的加速度,具体执行操作如下:
①降维。由于试验所获得数据是空间三维特征向量,无法在二维平面有效可视化,故通过PCA算法将原始数据转化为二维特征向量。
{A1,A2,A3,…,An}=PCA({X1,X2,X3,…,Xn}),
其中公式中Ai=[pi,qi]是降维后的二维特征向量。
②设置聚类中心个数。本文试验需要分析锦江牛的4类行为特征,所以将聚类中心设为4,分别为[α1,α2,α3,α4]。计算每个样本点到所有聚类中心的欧式距离,然后根据距离的大小将样本点归类到最近的中心所属的簇中。
D(i)=argmin‖Ai-αj‖2(j=1,2,3,4),
其中①中D(i)表示离样本Ai最近的聚类区域中心,αj为第j类的聚类中心。
③更新聚类中心。计算每个簇的特征均值zi,并将其作为该簇新的聚类中心。
其中n为所有样本的数量,I(·)为指示函数,当D(i)=j时,其值为1,否则为0,即将同属于j类别的样本相加然后求平均,也就得到新的聚类中心。
④迭代计算。重复执行③过程,不断对聚类中心进行迭代更新,直到聚类中心不再变化或者变化量小于设定的阈值。
通过上述4个算法步骤,最终可以将样本聚类成4个具有不同聚类中心(α1,α2,α3,α4)的簇,将收集的样本点分别与这4个聚类中心计算它们之间的欧氏距离,迭代计算后,将锦江牛的行为特征进行分类。
1.5 数据统计与分析
数据采用Excel 2010进行初步整理,SPSS 20.0软件的双因素统计方法分析采食、反刍、趴卧行为。采用Graph Pad 7.0软件绘图。
2 结果与分析
2.1 光照前期长光周期对锦江牛行为的影响
由表2双因素分析发现,光照前期LD组锦江牛站立时间比SD组降低35.1%(P<0.01),趴卧休息时间增加41.5%(P<0.01),趴卧反刍时间增加24.7%(P<0.05),采食时间降低11.1%(0.05 表2 光照前期长光周期对锦江牛行为的影响(n=18) min 长光周期对锦江牛趴卧率的影响见图1,LD组公牛的趴卧休息率在11:00显著提高(P<0.05),在13:00提高但差异不显著(0.05 由表3双因素分析发现,光照后期LD组锦江牛站立时间降低28.6%(P<0.01),趴卧时间提高20.9%(P<0.01);母牛趴卧反刍时间比公牛提高44.6%(P<0.01),采食时间增加22.3%(P<0.01)。光照与性别之间不存在互作效应。 由表4可知,四类行为特征中, LD组静止行为占比提高11.4%(P<0.05),其他行为特征差异不显著。 *表示差异显著(P<0.05),时间点为每次采血时间 表3 光照后期长光周期对锦江牛行为的影响(n=18) min 表4 长光周期对锦江公牛活动量的影响(n=9) % Phillips等[9]在研究光照对杂交小母牛影响时发现,光照对小母牛采食时间影响并不显著。Fuller等[4]的研究表明,长光周期(16L∶8D)与短光周期(8D∶16L)相比,小马驹的采食时间减少,而采食频率和干物质消化率增加,生长性能有所改善。Macmillan等[10]在探讨光周期管理对奶牛行为影响时,并没有发现光照周期对奶牛摄食行为有影响。邱静芸等[5]研究表明,锦江牛16 h光照组与自然光照组相比,采食时间显著降低。这与本试验LD组锦江牛采食时间有下降的趋势相一致,亦即长光周期使动物的采食形式发生改变,使采食的时间更加集中[11],肉牛的采食效率提高,表明食欲良好。 Oshiro等[12]研究表明,山羊白天反刍行为的发生频率显著高于夜间。Gordon等[13]发现,反刍行为的模式随所使用照明系统的改变而变化,并且不同的进料时间对反刍没有影响。Phillips等[14]探究奶牛对补光的差异反应时发现光照对奶牛反刍行为影响不显著。本试验中,光照对锦江牛总反刍行为时间影响不显著,但是,LD组静卧反刍时间在行为观察前期显著增加,在后期有增加趋势。此外,性别对采食时间具有显著影响,本次试验中母牛的采食时间长,可能是母牛体型较小,牛胃所能容纳饲料的体积也相对较小,因此母牛通过提高采食时间的方式摄入饲料。 本试验LD组锦江牛的趴卧休息时间极显著提高。Phillips等[14]在研究奶牛对补充光照的差异反应时发现,在奶牛趴卧区域补充光照会增加其趴卧时间。动物运动量的增加会加速营养消耗,因此,肉牛的趴卧休息率升高会减少其营养消耗,从而提高饲料的利用率,改善生产性能。韩兴泰等[15]在研究牦牛和黄牛不同运动量的能量代谢时发现,随着其运动速度的增加,能量消耗也在增加。本试验对锦江牛采食、趴卧、站立、反刍行为进行统计,LD组试验牛趴卧休息时间均显著高于12 h光照组,说明长光周期可以增加锦江牛的趴卧休息时间,减少能量消耗。 趴卧休息率是反映动物对其生活环境是否感到舒适的重要指标之一。趴卧休息率的增加能够有效反映畜牧场对动物生存环境的良好管理。Haley等[11]在探究奶牛在混凝土地板(不舒适环境)和床垫地板(舒适环境)的行为表现时发现,奶牛在舒适环境下会发生更多趴卧休息行为,但是采食与站立时间会减少,说明趴卧休息时间增加是动物表达对环境感到舒适的行为特征之一。趴卧时间又与动物的生长激素(GH)分泌密切相关,趴卧时间的减少会引起GH分泌减少,侧面反映趴卧休息率升高可提高动物的育肥效果。邱静芸等[5]探究长光照在不同时间点对肉牛趴卧率影响时发现,8:00、13:00、15:00肉牛的趴卧休息率显著上升。本试验分别探究7:00、9:00、11:00、13:00、17:00的趴卧状态,在11:00 LD组的趴卧休息率显著提高,13:00有提升趋势,母牛在9:00、13:00趴卧休息率也有增加的趋势,LD组锦江牛的趴卧率基本高于SD组,证实了长光照能提高动物的趴卧休息率。另外,邱静芸等[5]也证实采食率与趴卧休息率呈负相关关系,即趴卧休息率提升,食欲也随之改善。 Hanson等[16]通过三轴加速度传感器与GPS结合,不仅可以判定行为特征,更可以通过奶牛的行走距离来测定活动量水平。有研究通过直接观察与自动传感器收集奶牛行为表明传感器在探究动物行为中是一种有效的方法[17]。本试验采取活动量聚类分析方法剖析4种行为特征中,光照显著影响静止行为,LD组比SD组降低11.4%。 活动量是判定动物情绪是否安稳的重要指标之一。已有研究发现,奶牛处于发情焦虑状态下,其活动量会提升[18]。鄢新义等[19]在探究北京地区奶牛活动量时发现,奶牛的活动量在采食时达到最高峰,一年中夏季的活动略高于春秋两季,这些结果表明奶牛在采食兴奋或受到热应激情况下,奶牛的活动量会提升。奶牛的站立时间和行走时间长必然会引起活动量的提升,静止行为时间长证明奶牛活动量低,能量消耗少。此外,动物处于舒适环境下活动量会降低,不适的环境会引起动物不安,快走行为占比会上升[20]。本试验中,2组牛的快走行为占比几乎相等,说明长光照并不会引起锦江牛不适。 活动量可以作为研究动物神经活性的指标,动物神经兴奋性提高会导致动物活动量增加。本课题组曾报告妊娠后期活动量显著高于前期、中期,心率变异性分析表明后期低频成分(LF)显著高于前期、中期,可知后期交感神经兴奋性较高,导致奶牛后期活动量提高[21]。并且本研究LD组试验牛的静止行为特征比例上升,2组数据说明长光周期为锦江牛创造了舒适光环境,有利于改善肉牛的健康状态。 综上所述,长光周期增加锦江牛趴卧休息率并使反刍行为向趴卧反刍倾斜,增加静止行为状态时间,减少活动量有利于动物健康与生产。
2.2 光照前期长光周期对锦江牛趴卧率的影响
2.3 光照后期长光周期对锦江牛行为的影响
2.4 长光周期对锦江公牛活动量的影响



3 讨论
3.1 长光周期对锦江牛行为及趴卧率的影响
3.2 长光周期对锦江牛活动量的影响
猜你喜欢
农业科技通讯(2023年1期)2023-02-12上海文化(文化研究)(2021年5期)2021-11-12纺织科学研究(2021年9期)2021-10-14中国畜牧杂志(2021年8期)2021-08-15保健与生活(2019年15期)2019-09-12浙江农业学报(2019年7期)2019-08-14畜牧兽医学报(2018年7期)2018-08-07成都信息工程大学学报(2017年4期)2018-01-22少儿科学周刊·少年版(2017年7期)2017-09-29少儿科学周刊·儿童版(2017年7期)2017-09-29
