
利用90K芯片分析烟农74(11)小麦衍生品种(系)的遗传关系
2022-09-28 09:58翟晓灵王瑞霞张连焦娟李玉刚王圣健亓晓蕾盖红梅张胜全
中国农业科技导报 2022年8期
翟晓灵, 王瑞霞, 张连, 焦娟, 李玉刚, 王圣健, 亓晓蕾, 盖红梅*, 张胜全
(1.青岛市农业科学研究院, 山东青岛 266100;2.泰安市农业科学院, 山东泰安 271000;3.北京市农林科学院杂交小麦研究所, 北京 100097)
山东省烟台市农业科学研究院于1974年以丰产、抗病、落黄优良的洛夫林13为母本, 以71(17)6-1-1株系为父本配制杂交组合, 系统选育出烟农74(11)及74(11)混-1-1-3和C149等优良品系。烟农74(11)含有洛夫林13、蚰包麦、农林10号、北陆13、小罂粟(农林89)、欧柔的血统。洛夫林13来自罗马尼亚, 表现抗锈病和白粉病、耐高温;蚰包麦是当地的当家品种, 表现抗病、高产;农林10号、小罂粟来自日本, 携带Rht1、Rht2矮秆基因, 表现抗旱、矮秆、抗条锈病;欧柔来自智利, 表现大穗、大粒、抗条锈[1-2]。由烟农74(11)育成了20世纪90年代初大面积推广的鲁麦14、鲁麦13等品种, 2000年以后又育成了优质、高产品种烟农19。这些品种不仅在生产上发挥了很大的作用, 在小麦品种改良过程中也成为重要育种亲本, 衍生了济麦22、济南17、泰农18等大面积推广品种[3-5]。育种工作的突破性进展取决于关键遗传资源的发现与创新[6]。
SNP(single nucleotide polymorphisms)标记于1996年由Lander提出[7], 是生物基因组中普遍存在的单核苷酸多态性, 具有密度高、代表性强、遗传稳定性好和自动化程度高等优点, 得到专家学者的广泛关注。目前, 在不同物种中已探索出30多种SNP检测方法[8]。随着基因芯片和测序技术的快速发展,SNP标记已成为更加高效的第3代分子标记[9]。并在人类[10-11]、果蝇[12]、大麦[13]、玉米[14-15]等的遗传图谱构建、关联分析、QTL定位中发挥了重要作用。小麦基因组庞大, 其SNP研究滞后于水稻[16]、玉米[17]、大麦等物种[18]。2006年, 由Illumina公司研发的SNP GoldenGate芯片[19]大大推动了小麦基因组研究的发展, 目前, 小麦检测SNP常用的方法有重测序、小麦55K芯片、90K芯片、660K芯片等, 并在遗传多样性[20-21]、数量性状遗传[22-24]、全基因组关联分析[25-26]等方面得到广泛应用。
本研究利用90K SNP芯片对烟农74(11)的衍生品种(系)、亲本及近期育成小麦品种(系)进行了基因分型, 从系谱和分子水平探讨烟农74(11)种质在山东省小麦育种中的重要作用, 以期为小麦品种的改良提供参考。
1 材料与方法
1.1 供试材料
供试小麦品种(系)120份(表1), 其中1~62号为烟农74(11)的衍生品种(系), 编号63~120为非烟农74(11)衍生品种(系), 用于进行遗传多样性的对比分析。

表1 供试品种(系)Table 1 List of tested wheat variety(line) 续表Continued

表1 供试品种(系)Table 1 List of tested wheat variety(line)
1.2 研究方法
1.2.1 DNA提取 供试材料种植于田间, 在幼苗期剪取2片幼叶冷冻保存, 并标记取样单株。将冷冻样品用液氮研磨后, 用试剂盒(天根生化科技有限公司)提取基因组DNA[27], 并用NanoDrop Spectrophotometer(ND-1000)检测DNA样品浓度和质量。
1.2.2 小麦90K芯片分析和SNP分型 供试材料DNA的Illumina 90K芯片分析由北京博奥晶典生物技术有限公司完成。采用Genomestudiov2011.1软件包对结果文件*.bmp和*.idat进行SNP数据校正和分型[28]。数据校正基于芯片的原始数据, 即Genomestudiov2011.1软件生成的每个SNP位点的聚类图, 如果聚类图分为2个组群(无杂合位点)或3个组群(有杂合位点), 而且每组表现为紧密簇拥在一起, 则这些聚类图表示的SNP位点为有效SNP位点, 可用于供试材料的遗传多样性分析;如果聚类图分为杂乱无章的多个组群, 则判定为无效SNP位点, 不能用于供试材料研究, 需在Genomestudiov2011.1软件中, 将无效SNP位点标记出来, 待全部位点全部校正完, 导出SNP基因型数据, 将其中标记无效的位点去掉, 即完成全部SNP位点校正。
1.2.3 遗传多样性分析 利用Excel软件进行SNP分型数据的整理和存储, 采用自主研发程序将原始SNP数据的AA、AB、BB基因型转换为Powermarker3.25软件[29]需要的输入格式。利用PowerMarker 3.25软件分析多态性信息含量(polymorphism information content, PIC)、基因多样性、每个SNP位点的等位变异数、主效等位变异频率、期望杂合度及遗传距离, 并计算遗传相似性系数。其中, PIC、基因多样性等用于评估群体的遗传多样性, 遗传相似性系数用于评估2个供试材料之间的遗传关系。

2 结果与分析
2.1 烟农74(11)种质对山东省冬小麦品种的支撑作用
由图1可看出, 以烟农74(11)种质为亲本, 选育出了鲁麦14、鲁麦13、烟农19等大面积推广品种。这些品种不仅在小麦生产中大面积推广应用, 还在小麦育种中发挥了重要作用。
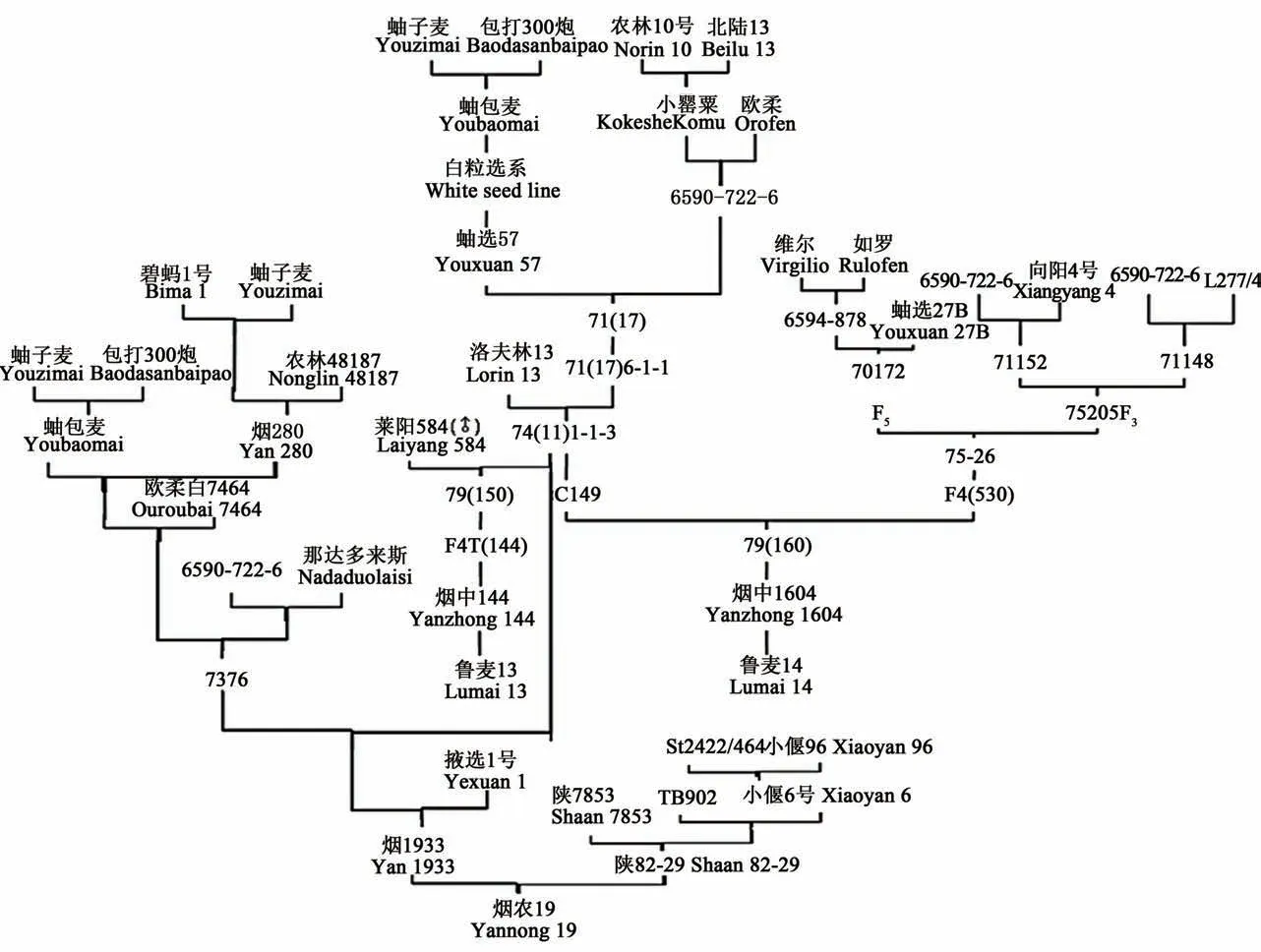
图1 小麦关键种质烟农74(11)的系谱追踪及主要衍生品种Fig.1 Pedigree tracking of 74(11)and the principal derived variety
对2000—2020年山东省审定小麦品种进行了系谱溯源, 21世纪以来20年间共审定小麦品种197个, 有55.33%的品种含有烟农74(11)种质血统, 36.04%的品种含有鲁麦14血统, 21.32%的品种含有鲁麦13血统, 10.66%含有济麦22血统, 4.57%含有烟农19血统。由图2看出, 不同年代材料在育种中发挥的作用不同, 就烟农74(11)种质而言, 在国家“十五”到“十三五”计划期间, 其直接衍生的品种占比分别为28.57%(36个品种)、66.67%(36个品种)、62.07%(29个品种)、58.76%(97个品种), 表明该种质对山东省小麦新选育品种具有系谱支撑作用。此外, 鲁麦14作为烟农74(11)种质的直接衍生品种, 在4个五年计划中的衍生品种占比分别为20.00%, 41.67%、37.93%和39.18%, 表明鲁麦14在小麦育种中成为重要的育种亲本。
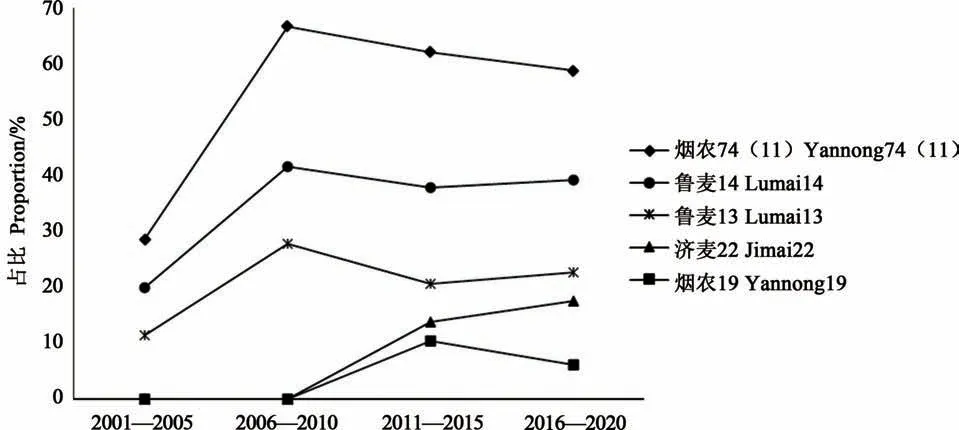
图2 2000—2020年烟农74(11)及其衍生品种(系)在山东省的衍生材料统计Fig.2 Statistics of wheat materials derived from Yannong 74(11)and the principal variety(line)inShandong province from 2000 to 2020
2.2 供试材料的SNP分型及遗传多样性分析
120份供试材料共获得有杂交信号的SNP位点81 587个, 校正后获得有效SNP位点26 026个, 有染色体定位的SNP位点20 953个, 其中A基因组8 373个, B基 因 组10 592个, D基 因 组1 988个, 覆盖小麦基因组3 646.01 cM。
小麦90K芯片获得的SNP基因型数据在染色体间分布非常不均匀, 变异系数为58.17%, SNP个数最少的染色体是4D(73个), 最多的是1B(1 984个), 且在3个基因组中, 均为D基因组的SNP个数最少。SNP标记的平均等位变异数为2.28个, 变异系数为1.80%, 变幅很小。平均主效等位基因频率为0.77, 平均基因多样性为0.31, 平均期望杂合度为0.03。平均多态性信息含 量(PIC)为0.25, 变 幅 为0.18(4D)~0.31(2D), 明显低于SSR标记揭示的遗传多样性(表2)。
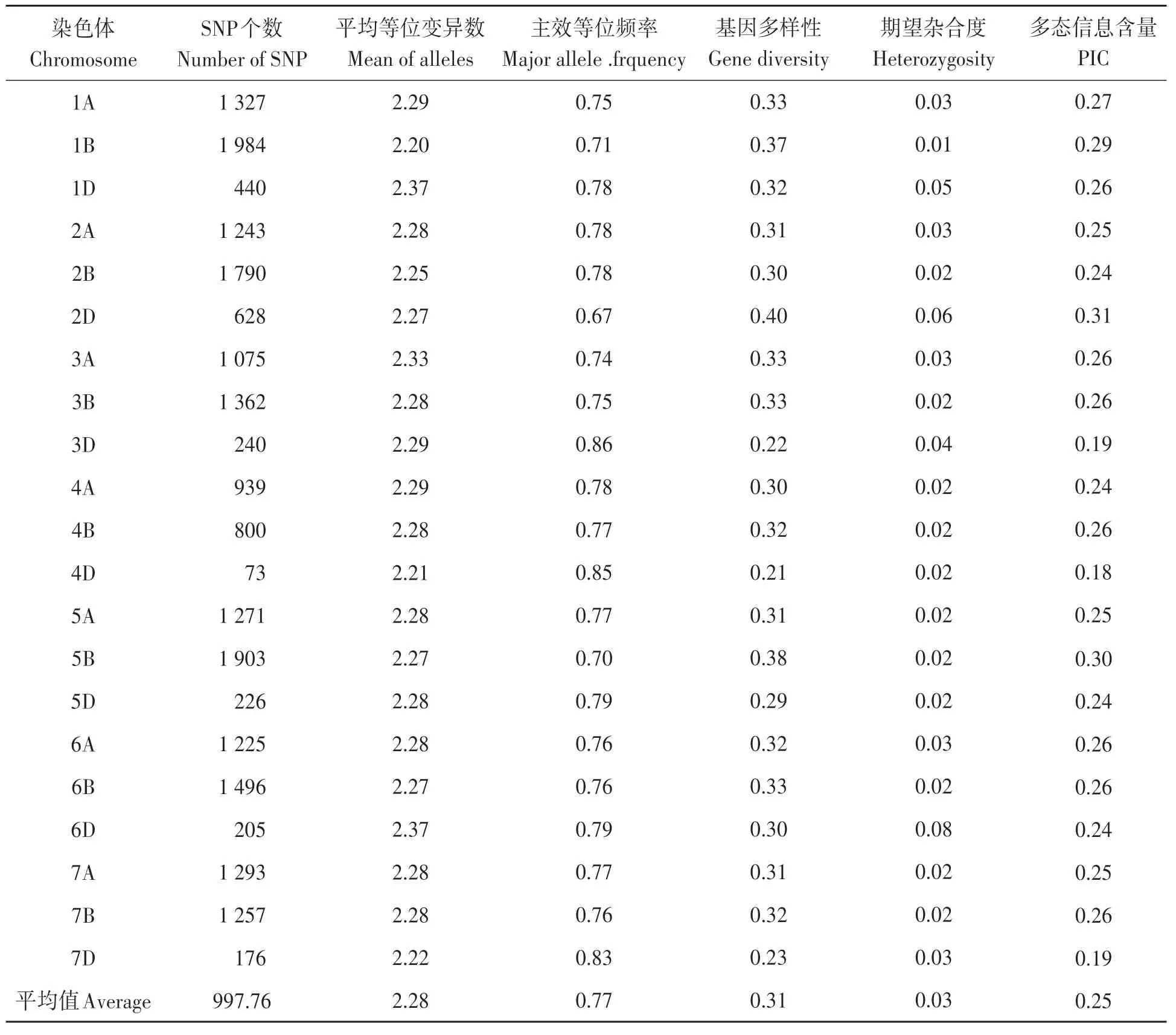
表2 供试材料在染色体水平的遗传多样性Table 2 Genetic diversity of SNP marker at chromosome level with tested materials
2.3 不同供试材料分组的遗传关系
对供试材料进行了分组, 分为全部材料组(组Ⅰ)和烟农74(11)种质衍生材料组(组Ⅱ)(图3)。对2组供试材料分别进行了遗传相似性系数的计算, 组Ⅰ共有7 140个两两组合模式, 遗传相似性系数范围为0.56~1.00, 平均为0.70;根据遗传相似性系数区间对材料进行了统计分析:0.51~0.60的 材 料 占0.66%、0.61~0.70的 占59.44%、0.71~0.80的占32.98%、0.81~0.90的占6.12%、≥0.91的占0.80%, 近60%的材料间遗传相似性系数在0.61~0.70区间, 表明组Ⅰ材料的遗传关系相对较远。组Ⅱ材料之间共有2 219个两两组合模式, 遗传相似性系数范围为0.61~1.00, 平均为0.75;遗传相似性系数区间在0.61~0.70的材料占26.77%、0.71~0.80的占53.22%、0.81~0.90的占17.26%、≥0.91的占2.75%, 表明仅含有烟农74(11)种质衍生后代之间的遗传相似性系数一半以上为0.71~0.80, 近20%的材料组合为0.81~0.90, 表明组Ⅱ的材料两两间遗传关系比组Ⅰ更近。

图3 供试材料两两之间的遗传相似性系数统计Fig.3 Statistical of genetic identity coefficient between every two tested materials
2.4 典型组合遗传关系
烟农74(11)种质衍生后代中系谱关系比较完整且在小麦生产中大面积应用的小麦品种与亲本之间的遗传关系如表3所示。主要杂交组合为:鲁麦13/临汾5064、鲁麦14/豫麦13、鲁麦14/鲁884187、鲁麦13/豫麦2号。由于济麦22的父本材料未得到妥善保存, 故只分析了济麦22与其母本935106的遗传关系。分析发现, 这4个杂交组合的父母本之间的遗传相似性系数为0.69、0.65、0.68、0.71, 平均为0.68;子代与上缘亲本间的遗传相似性系数为0.67~0.87, 平均为0.79;姊妹系之间的遗传相似性系数为0.84~0.87, 平均为0.855。济麦22与母本的遗传相似性系数为0.87, 与骨干亲本鲁麦14的遗传相似性系数为0.85。从父母本之间的遗传关系可以看出, 这些组合双亲的遗传关系相对较远;从子代与亲本的遗传关系可以看出育种中存在偏向性选择现象, 其中子代与关键亲本的遗传相似性系数达0.80以上, 甚至0.85以上。

表3 基于小麦90K芯片的典型杂交组合亲本及后代的遗传关系Table 3 Genetic relationship of typical test material based on wheat 90K array 续表Continued
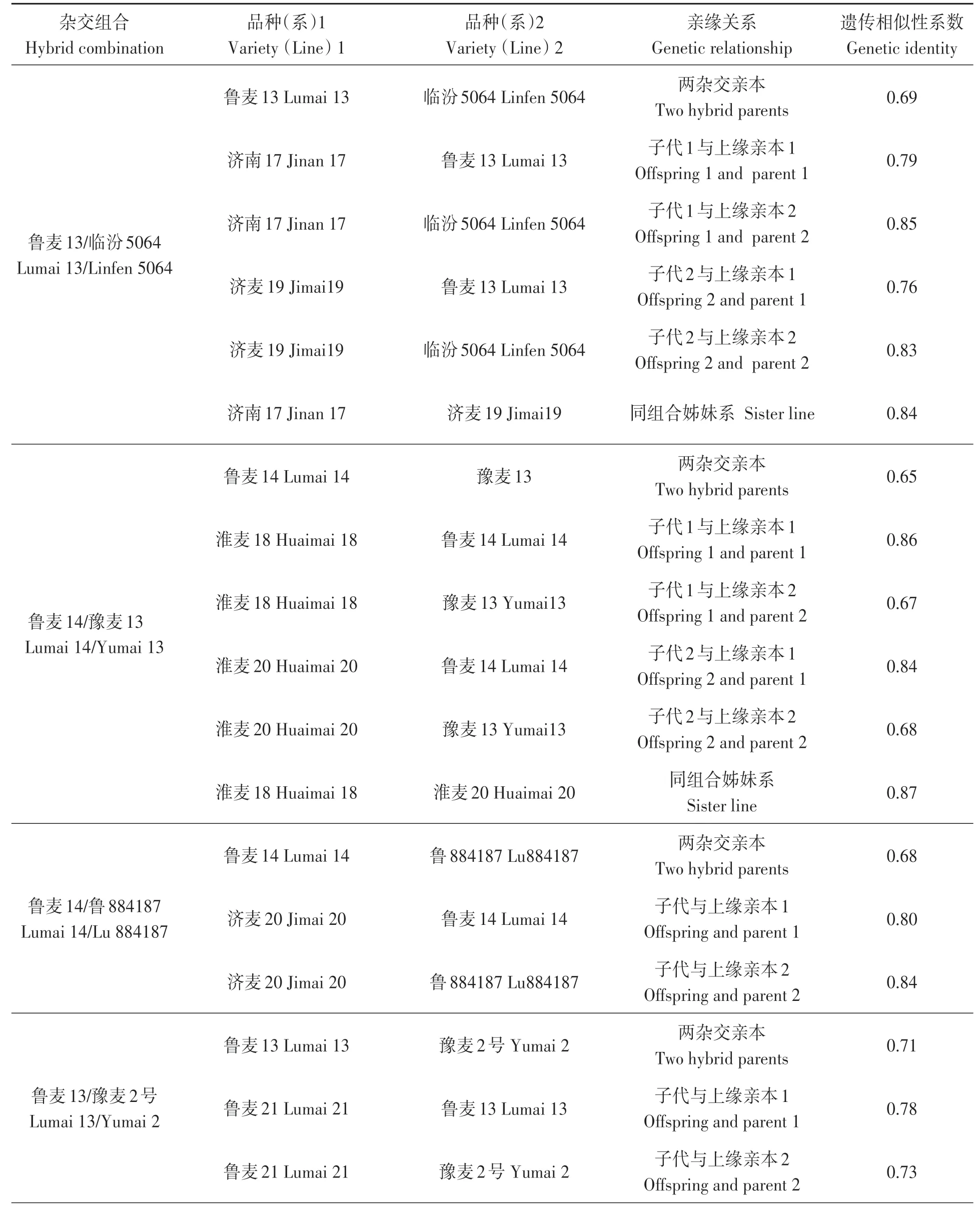
表3 基于小麦90K芯片的典型杂交组合亲本及后代的遗传关系Table 3 Genetic relationship of typical test material based on wheat 90K array
3 讨论
本文对120份供试小麦材料进行了90K芯片分析, 结果表明,这些供试材料的平均遗传相似性系数为0.70, 如果排除蚰包麦、农林10号等早代育种亲本, 则当代育成品种(系)的遗传关系更近:62份烟农74(11)种质的衍生后代间的平均遗传相似性系数为0.75。当前, 极端灾害天气频发, 病虫害日益加重, 遗传种质的单一化, 容易导致抗灾能力减弱, 甚至有“全军覆没”的风险。刘建军等[30]认为, 由于长期对产量、抗性、品质等性状的持续定向选择, 导致了当代育成品种遗传多样性急剧下降。遗传多样性的降低在一定程度上阻碍了小麦突破性品种的选育, 从而阻碍小麦育种的发展和潜力的提高。李邦发[31]指出, 繁6及其衍生品种在四川占统治地位30年, 但由于2001年条锈病大流行, 超过90%的衍生品种感染条锈病, 造成小麦生产巨大损失, 在育种领域应引以为戒。因此, 应扩大小麦育种材料的遗传基础, 寻找更加多样的优良骨干亲本。
烟农74(11)种质来自20世纪50、60年代的蚰子麦衍生的蚰包麦, 其在山东省连续推广种植15年, 是黄淮麦区首个产量超过7 500 kg·hm-2的小麦品种[5], 被庄巧生先生列为16个骨干亲本之一[1]。张学勇等[32]认为, 早期的品种或骨干亲本无论当时多么优秀, 其田间表现和遗传背景与今天的优良品种或骨干亲本相比都有很大差距, 这反映了小麦品种改良是早期品种的继承和符合现代目标的优良基因积累的过程。蚰子麦→蚰包麦→烟农74(11)种质→鲁麦14→济麦22的衍变过程, 也表明不同年代的当家品种首先是在继承和综合中发展, 必需有经过生产检验的品种基因组为基础;此外, 从一代当家品种发展到新一代当家品种, 必需有突破性的进展, 必需解决当家品种与生产实际之间的主要矛盾。
致谢:感谢烟台市农业科学研究院方正先生在烟农74(11)种质的特征特性及育种地位等方面给予的指导。
猜你喜欢
农村科学实验(2022年15期)2022-10-13
——以韶关市为例
安徽农业科学(2022年14期)2022-08-04
西南农业学报(2022年5期)2022-06-06
热带亚热带植物学报(2022年2期)2022-04-14
三农资讯半月报(2020年13期)2020-07-31
计算机技术与发展(2019年1期)2019-01-19
雪莲(2017年2期)2017-05-12
环球市场信息导报(2017年1期)2017-04-08
江苏农业科学(2016年4期)2016-06-14
源流(2016年8期)2016-05-14
