
陆地棉果枝夹角性状的主基因+多基因混合遗传模型分析
2023-01-05 08:45马麒李吉莲徐守振陈红刘文豪宁新柱林海
生物技术通报 2022年10期
马麒 李吉莲 徐守振 陈红 刘文豪 宁新柱 林海
(新疆农垦科学院棉花研究所 农业农村部西北内陆区棉花生物学与遗传育种重点实验室 新疆兵团棉花改良与高产栽培重点实验室,石河子 832000)
棉花(Gossypium spp.)不仅是纺织工业重要的天然纤维来源,而且是关系国计民生的重要战略物资,在我国国民经济中的作用举足轻重[1]。新疆是我国最大的棉花生产基地,也是植棉机械化程度最高的地区。随着新疆“矮、密、早、膜”栽培模式和机采棉的大面积推广,合理株型结构在棉花生产中的作用日益凸显。
陆地棉(Gossypium hirsutum)是世界上种植面积最大的四倍体栽培种。在陆地棉株型相关性状中,果枝夹角不仅决定着棉花的花器官和棉铃的空间分布,而且还决定着植株整体框架结构的紧凑程度,是适宜机采棉花的株型结构中最关键的因子之一。一般来讲,相对较小的果枝夹角会使得棉花株型结构较为紧凑,能有效提高冠层透光性,塑造出相对合理的株型,更适宜合理密植、轻简化栽培及机械采摘,达到增产增效的目的。因此,果枝夹角小已经成为当前棉花的重要育种目标性状之一。长期以来,陆地棉作为一种重要的纤维作物,人们十分关注其纤维产量相关性状,如单铃重、衣分、单株结铃数等[2-3],以及品质相关性状指标,如纤维上半部平均长度、断裂比强度、马克隆值等[4-5],很少有人对其果枝夹角性状进行深入测定、解析和研究。陆地棉果枝夹角性状是典型的数量性状,受基因型和环境共同控制和影响[6-8],明确果枝夹角的遗传特性对提高棉花株型育种效率具有重要意义。
研究表明,植物数量性状的通用性模型为主基因+多基因混合遗传模型,单纯主基因或多基因模型均为其特例[9]。李成奇等[10]也认为植物数量性状的遗传特征不仅是一种多基因遗传模式,还存在主基因+多基因混合遗传模式。盖钧镒等[11]认为该模型不仅可以检测和鉴定数量性状主基因和多基因的存在,还能估测其遗传参数,可作为作物数量性状遗传解析的重要方法之一。近年来,该遗传分析方法已在水稻[12-13]、小麦[14-15]、花生[16-18]、玉米[19-20]、油菜[21-22]、甜瓜[23]、蓖麻[24]、胡麻[25]等多种作物中得到广泛应用。在棉花上主要应用于棉花的早熟性[26]、生育期[27-28]、产量[29-30]、品质[29]、黄萎病[31]、衣分[32]、铃重[33-34]、抗逆性[35]以及叶柄分化率[36]等性状的遗传分析,而在果枝夹角性状遗传解析方面的研究相对较少。张培通等[37]利用陆地棉泗棉3号和CARMEN构建 RIL群体,利用P1、P2和RIL群体三世代联合分析法对棉花株型性状进行了主基因+多基因遗传分析,结果表明,果枝夹角性状符合主基因+多基因混合遗传模型,并且存在主基因,但其主基因遗传率较低;而李成奇等[10]对陆地棉品种百棉 1号主要株型性状进行主基因+多基因混合遗传分析发现,果枝夹角遗传表现为多基因遗传或以多基因遗传为主,不存在主基因。
为进一步探明陆地棉果枝夹角的遗传特性和规律,本研究首先以不同来源的陆地棉种质资源组成的自然群体为研究对象,研究果枝夹角在不同生态环境下的表型值,从中筛选出果枝夹角极端差异材料,作为亲本构建四世代联合群体(P1、P2、F1、F2),采用植物数量性状主基因+多基因混合遗传模型方法,对4个世代群体的果枝夹角表型性状开展多世代联合遗传分析,探讨多世代陆地棉果枝夹角性状的遗传规律,并估测主基因遗传效应与遗传率,从而为陆地棉果枝夹角性状的分子遗传解析和棉花株型育种提供理论依据和参考信息。
1 材料与方法
1.1 材料
试验材料主要是由418份陆地棉种质资源组成的自然群体,以及从中筛选出来的果枝夹角极端差异的两个亲本材料中2549和新陆早18号。418份陆地棉种质包括前苏联材料27份,美国材料57份,我国北方特早熟棉区材料14份,西北内陆棉区材料28份,长江流域棉区材料107份,黄河流域棉区材料167份。亲本材料P1(中2549)为果枝夹角较大的材料作父本,P2(新陆早18号)为果枝夹角较小的材料作母本(图1),中2549来自于黄河流域棉区,新陆早18号为新疆自育陆地棉品种。2020年4月分别在新疆农垦科学院棉花研究所石河子试验地种植,以中2549为父本,新陆早18号为母本配制杂交组合,得到F1种子,同年10 月在海南种植F1群体,自交获得F2种子。2021 年4 月分别在新疆农垦科学院棉花研究所石河子试验地和铁门关试验地种植P1、P2、F1和F2群体,并进行果枝夹角表型性状鉴定。

图1 果枝夹角极端差异亲本单株材料Fig.1 Parents with extremely different FBA
1.2 方法
1.2.1 田间种植方式 田间材料的种植方式均采用新疆普遍推广的66 cm+10 cm的机采棉株行距配置模式,平均行距为38 cm,石河子种植模式为一膜6行,一穴一株,株距9.5 cm,膜宽为2.05 m;铁门关和阿拉尔的种植模式为一膜4行,一穴一株,株距9.5 cm,膜宽为1.25 m。敦煌种植模式为一膜3行,行距40 cm,株距15 cm,膜宽1.2 m。田间管理均按照当地标准大田。
1.2.2 性状调查 果枝夹角性状的鉴定是在吐絮期进行的。自然群体测定时每份材料选取连续相邻的10株为调查对象;对亲本P1、P2,以及F1和F2群体的果枝夹角性状进行调查测定时,P1、P2各调查80 个单株,F1调查30个单株,F2调查200 个单株。具体测定方法是:利用捷立(GeeLii)数显角度测量仪对棉株中部果枝(基部起第4、5、6台)夹角进行调查鉴定,并取其平均值。
1.2.3 遗传模型分析 采用盖钧镒等[38]发明的主基因+多基因混合遗传模型分析方法,对P1、P2、F1和F2共4 世代联合分析果枝夹角性状的遗传规律和基因互作方式。通过比较由1 对主基因(1MG)、2 对主基因(2MG)、多基因(PG)、1对主基因+多基因(MX1)、2对主基因 + 多基因(MX2)组成的共24 个遗传模型的AIC 值(Akaike′s information criterion),进行遗传模型的适合性检验(均匀性检验、Smirnov 检验和Kolmogorow 检验,获得5 个基本统计量U12、U22、U23、nW2和 Dn)来选择最优模型。根据模型分析结果,利用最小二乘法估计入选模型的主基因和多基因效应值、方差及相关遗传参数。该分析过程主要通过混合遗传分析 R 软件包SEA 程序[39]。
2 结果
2.1 陆地棉主茎不同部位果枝夹角的表型差异性分析
在前期的预试验中,从418份陆地棉核心种质资源中各选取代表性材料30份,分别在新疆石河子、库尔勒两个生态试验点测量棉花整株所有果枝的夹角大小,并开展不同节位的果枝夹角差异性分析。结果表明,陆地棉植株整株的果枝夹角大小整体表现出如下特征:(1)从植株基部到顶部果枝夹角呈现出整体下降的趋势,第1-9台果枝夹角大小分别为 59.22°±1.35°、57.21°±2.14°、57.18°±1.09°、56.02°±0.94°、55.94°±1.98°、55.19°±1.15°、54.47°±2.31°、52.59°±1.92°、54.05°±1.12°(图2);(2)下部果枝夹角(1-FBA,基部起第1-3台果枝)、中部果枝夹角(2-FBA,基部起第4-6台果枝)和上部果枝夹角(3-FBA,基部起第7-9台果枝)表现出极显著(P<0.01)的差异性,但 2-FBA与整株的果枝夹角(All-FBA)之间无显著差异性(P>0.05)(表1)。相关性分析表明,1-FBA、2-FBA、3-FBA均与 All-FBA 呈极显著(P>0.01)的正相关性,Pearson 相关系数均大于0.50。此外,2-FBA 与 All-FBA 的相关性最强,Pearson 相关系数为 0.71(表2)。上述研究结果表明,陆地棉中部果枝(2-FBA)夹角与整株果枝夹角的相关性最强,且二者的果枝夹角差异性最小,所以中部果枝夹角(第4-6台)最能够代表陆地棉整株的果枝夹角。这一研究结果为后续试验研究的开展奠定了重要基础。

表2 各部位果枝夹角的相关性分析Table 2 Correlation analysis of FBA in different positions

图2 不同果枝部位果枝夹角变化趋势图Fig.2 Variation trend of FBA in different fruit branch position

表1 不同部位果枝夹角的差异性比较Table 1 Comparison of FBA in different positions
2.2 陆地棉自然群体果枝夹角的表型统计分析
通过对自然群体在4个环境下果枝夹角性状的鉴定与分析,结果发现418份陆地棉种质资源果枝夹角在4个种植环境下具有广泛的变异,单个环境下果枝夹角的极差范围是14.99°-24.67°;各环境表型的平均值范围是(56.15°±4.13°)-(62.61°±2.61°)。整体来看,果枝夹角的变异度相对较小,各环境下的变异度范围是4.17%-7.36%,并且不同环境下果枝夹角的变异度相对比较稳定。在4个环境中,石河子的平均果枝夹角最大,为62.61°±2.61°,但变异度却最小,为4.17%;而阿拉尔的平均果枝夹角最小,为56.15°±4.13°,但变异度最大,为7.36%。4个环境下果枝夹角的平均值为58.85°±3.30°,平均变异度为5.63%。(表3)通过柯尔莫哥洛夫-斯米诺夫(Kolmogorov-Smirnov,K-S)正态分布检验发现,在E1、E2、E3、E4、Eall环境下,Z值分别为0.557、0.734、0.913、0.646、0.744,P值分别为0.916、0.654、0.375、0.789、0.637,P 值均大于 0.05(表3)。频率分布直方图和P-P图显示,陆地棉果枝夹角在E1、E2、E3、E4、Eall环境下均呈现出典型的正态分布趋势(图3),说明陆地棉果枝夹角在4个单一环境和总环境下均表现出显著的正态分布,符合一般数量性状的遗传特点。
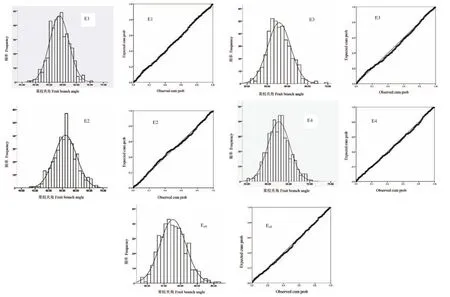
图3 果枝夹角性状在不同种植环境下的正态分布图Fig.3 Normal distribution of FBA trait under different planting environments

表3 陆地棉自然群体果枝夹角在4个环境下的表型统计分析Table 3 Phenotypic statistical analysis of FBA of G.hirsutum under four environments
2.3 各世代群体果枝夹角性状的表型频次分布
由表4可知,各世代 P1、P2、F1、F3群体的果枝夹角的平均值分别为 64.05°、49.57°、56.86°、52.15°,极差分别为 11.79°、38.78°、34.01°、54.40°(表4)。对各世代群体的果枝夹角表型数据进行正态分布分析,结果发现除P2群体外,其它各世代群体的果枝夹角数据均呈现出连续的正态分布(图4)。通过K-S正态分布检验发现,各世代群体P1、P2、F1、F3的K-S Z值分别为0.85、2.23、0.58、1.04,其对应的渐近显著性P值分别为0.46、0.00、0.88、0.23(表5)。可以看出,只有P2的渐近显著性P值小于0.05,其它各个世代群体均大于0.05。上述结果表明,果枝夹角性状属典型的数量性状,符合数量性状遗传的一般特点。

表5 各世代群体果枝夹角性状表型数据的K-S正态分布检验Table 5 Kolmogorov-Smirnov test of phenotypic data of FBA in different generations
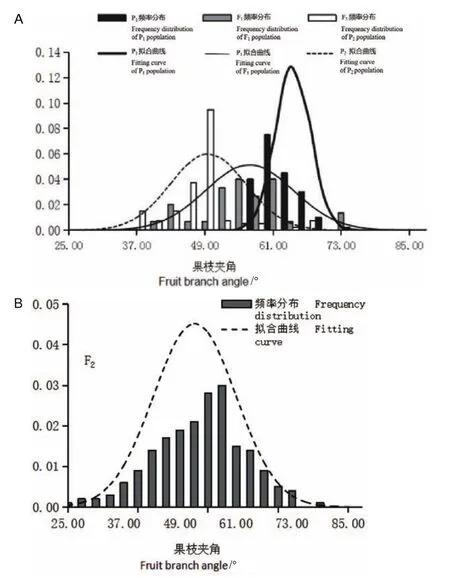
图4 各世代群体果枝夹角的正态分布图Fig.4 Normal distribution of FBA for each generation population
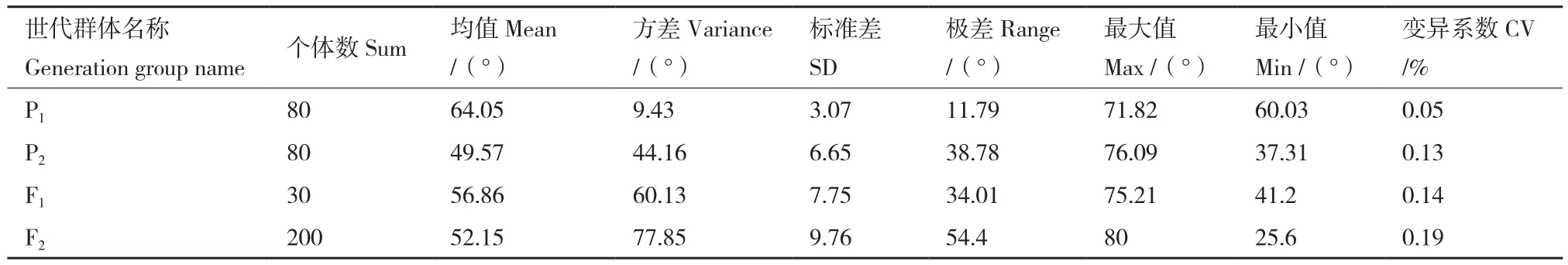
表4 各世代群体果枝夹角性状的表型数据分布Table 4 Phenotypic data distribution of FBA in each generation population
2.4 果枝夹角的主基因+多基因遗传模型分析
2.4.1 果枝夹角的最适遗传模型的选择 采用植物数量性状主基因+多基因混合遗传模型的多世代联合分析方法,对中2549 和新陆早18号组合的4 个世代群体的果枝夹角进行遗传分析,获得24 种遗传模型的极大似然函数值,并根据极大似然值计算出遗传模型的AIC值(表6)。通过比较24个遗传模型的AIC值,从中选择出AIC值最小或接近的遗传模型作为候选遗传模型。比较发现,1MG-EAD(1对等加性-显性主基因)、1MG-NCD(1对负向完全显性主基因)、2MG-EA(2对等加性主基因)、2MGEAD(2对等加性-显性主基因)模型的AIC值相对较小,分别为2 626.710、2 626.977、2 624.698、2 624.816,可作为果枝夹角的候选遗传模型。

表6 24个不同遗传模型的极大似然值(MLV)及AIC值Table 6 Max-likelihood-value(MLV)and Akaike’s information criterion(AIC)of 24 different genetic models
进一步分别对每个候选遗传模型进行适合性检验,包括均匀性检验(U12、U22、U32)、Simironow 检验(nW2)和Klmogorov检验(Dn)。并选择适合性检验统计量达到显著水平个数最少且AIC最小的模型作为最适模型。由表7可知,4个候选遗传模型的适合性检验统计量达到显著性水平的个数相等,所以选择AIC值最小的模型2MG-EA作为果枝夹角的最适遗传模型,即2对等加性主基因模型,表明陆地棉果枝夹角存在主基因遗传。

表7 果枝夹角候选遗传模型的适合性检验Table 7 Fitness test for candidate genetic models for FBA
2.4.2 最优遗传模型的遗传参数估计 根据果枝夹角遗传模型的极大似然估计值,估计最适遗传模型的遗传参数。在确定出最佳遗传模型的条件下,采用最小二乘法,通过成分分布参数估计主基因的遗传参数(表8)。从表8中可知,控制果枝夹角的两对等加性主基因的加性效应值为3.65,主基因遗传率为90.22%。这说明陆地棉果枝夹角性状主要受主基因控制,且主基因遗传率较高。

表8 果枝夹角最优遗传模型的遗传参数估计Table 8 Estimation of genetic parameters of FBA under its optimal genetic model
3 讨论
在农作物上,分枝角度是决定作物株型的重要农艺性状之一。分枝角大的植株会占用过多空间,降低作物的种植密度,在生长阶段的后期增加阴影面积和倒伏情况,从而降低产量[40]。相反,相对较小的分枝角度是作物合理密植的前提和获得较高产量的保障。譬如,在长期的品种选择和遗传改良过程中,随着水稻分枝角度的减小,水稻由松散株型的野生稻改良成紧凑株型的栽培稻,其耐密性和光合效率显著提升,从而有效实现产量提高[41]。对棉花来说,小果枝夹角塑造了棉花相对紧凑的株型结构,有助于提高其机械化收获效率。例如,在相同的种植条件下,相对较小果枝夹角的棉花有利于其群体冠层的透光性,便于吐絮期各部位叶片的脱落,从而有效降低机采棉的籽棉含杂率,显著提高机采棉的原棉品质[42-43],从而达到提质增效的目的。综合来看,相对较小的分枝角度在农作物生产上更有利于其合理密植、机械化采收及产量的提高。
尽管植物分枝在一定程度上也受栽培模式、化学调控和外部生长环境等因素的影响,但植物本身的遗传特性是决定分枝角度的主要因子[44-45]。因此,阐明陆地棉果枝夹角的遗传规律对于其株型结构育种具有重要的理论指导意义。本研究采用主基因+多基因遗传分析方法,系统地对果枝夹角性状进行了遗传规律的解析。结果表明,果枝夹角检测到两对等加性的主基因,通过遗传参数估计,主基因遗传率为90.22%,这说明陆地棉果枝夹角性状主要受主基因控制,且主基因遗传率较高。进一步表明该性状的遗传变异主基因作用大于多基因,对其可采用简单杂交重组或回交转育的方法即可实现目标性状主基因的转入,同时兼顾增效多基因的聚合[10],并且在早期世代进行果枝夹角的选择是行之有效的方法[46]。但就关于果枝夹角性状遗传特性的研究,前人的研究结果不尽一致。张培通等[36]利用P1、P2和RIL群体三世代联合分析法对棉花株型性状进行主基因+多基因遗传分析,研究发现果枝夹角性状符合主基因+多基因混合遗传模型,同样认为存在主基因,其研究结果与本文一致,但与本文不一致的是认为主基因遗传率较低。而李成奇等[10]通过对百棉 1号主要株型性状进行主基因+多基因混合遗传分析,发现果枝夹角遗传表现为多基因遗传或以多基因遗传为主,不存在主基因。这说明本研究结果与前人的研究结果也不尽一致,这可能与研究材料、性状鉴定方法等不同有关。
另外,本研究选取的陆地棉果枝夹角极端差异的亲本材料,是从418份陆地棉种质资源构成的自然群体中筛选鉴定的,因此在果枝夹角极端差异性方面显著性较强,研究材料的选取上更具有代表性。其次,在果枝夹角性状的鉴定方面,本研究通过开展陆地棉主茎不同部位果枝夹角与整株果枝夹角之间的相关性、差异显著性分析,发现中部果枝(第4-6台)夹角最能够代表陆地棉整株的果枝夹角,提出以中部果枝夹角作为陆地棉果枝夹角性状鉴定的目标参照,减少了果枝夹角性状调查的工作量,调查方法具有一定的创新性。此外,在果枝夹角性状鉴定过程中,本研究采用的主要测量工具是数显角度测量仪,该角度测量仪器具有电子感应、操作简单、测量精准的优点,为本研究的顺利进行提供了工具保障,这与前人的调查方法也不尽相同。本研究结果有助于阐明陆地棉果枝夹角性状的遗传规律,对于陆地棉果枝夹角的分子遗传解析及株型遗传育种具有重要现实意义。
4 结论
陆地棉果枝夹角变异度相对较小,中部果枝(基部起第4-6台)夹角最能够代表陆地棉整株的果枝夹角水平。控制果枝夹角性状的最佳模型为2对等加性主基因模型,主基因的加性效应值为3.65,遗传率为90.22%。这说明陆地棉果枝夹角性状主要受主基因控制,且主基因遗传率较高。
猜你喜欢
中国棉花(2022年6期)2022-08-16
农业科技通讯(2022年6期)2022-06-21
作物学报(2022年7期)2022-05-11
中学生数理化·七年级数学人教版(2020年12期)2021-01-18
语数外学习·高中版上旬(2020年8期)2020-09-10
新疆农业科学(2020年2期)2020-03-12
中学生数理化·高一版(2018年5期)2018-06-04
新高考·高二数学(2017年8期)2018-03-13
中国农业信息(2017年12期)2017-09-03
养生保健指南(2016年12期)2017-01-06
