
Nrf2和IGF-1在胃黏膜癌变过程中的表达
2012-11-21 01:12宋素贞王洪波山东中医药大学第二附属医院职业病科山东济南25004
中国老年学杂志 2012年17期
宋素贞 王洪波 (山东中医药大学第二附属医院职业病科,山东 济南 25004)
Nrf2和IGF-1在胃黏膜癌变过程中的表达
宋素贞 王洪波1(山东中医药大学第二附属医院职业病科,山东 济南 250014)
目的 研究核因子相关因子-2(Nrf2)及胰岛素样生长因子-1(IGF-1)在胃正常黏膜、增生性息肉、上皮内肿瘤(GIN)及腺癌中的表达情况,并初步探讨二者在胃癌癌变过程中的作用机制。方法 应用免疫组化方法检测Nrf2和IGF-1在胃正常黏膜、增生性息肉、各级GIN及不同分化程度腺癌中的表达情况。结果 Nrf2和IGF-1在正常胃黏膜中几乎无表达;从增生性息肉、GIN到腺癌,二者共表达逐渐增强;在增生性息肉、GIN、腺癌中的表达差异有统计学意义(均P<0.05),并且呈正相关(r=0.337,P=0.037)。结论 Nrf2和IGF-1表达增强可能与胃黏膜癌变过程有关,二者表达水平升高有可能促进肿瘤进展。
核因子相关因子-2;胰岛素样生长因子-1;胃癌;致癌作用;氧化应激
胃癌前病变在转变为胃癌的过程中,由于受到亚硝酸盐、酒精、胆汁反流及幽门螺杆菌(HP)感染等外源性氧化应激源的刺激,从而引起氧化应激反应,致使机体组织细胞中产生过多活性氧自由基,导致细胞DNA损伤及突变,从而导致胃癌的发生。核因子相关因子-2(Nrf2)与机体的防御系统密切相关,是细胞防御氧化应激的重要调节因子,通过介导一系列抗氧化酶和Ⅱ相解毒酶的表达,可以保护多种组织免受有害物质的攻击。因此,Nrf2已被列为肿瘤化学预防候选基因〔1,2〕。在既往研究中发现Nrf2基因敲除鼠恶性肿瘤的发病率明显高于野生鼠。胰岛素样生长因子-1(IGF-1)有显著的抗凋亡作用,对肿瘤细胞的耐药性起关键作用〔3,4〕。本研究拟探讨Nrf2及IGF-1在胃癌癌变过程中的作用机制。
1 材料与方法
1.1 材料 本研究中283例病人胃组织标本来源于山东大学第二医院。其中,腺癌135例,来自2001~2005年病理科手术切除标本;上皮内肿瘤(GIN)96例(62例低级别以及34例高级别,包括6例原位癌)、增生性息肉30例及正常胃黏膜组织22例,均来自2005~2008年胃镜室活检标本。临床及病理信息包括姓名、性别、年龄、肿瘤浸润深度、组织类型及淋巴结转移情况等均来自临床资料。Nrf2、IGF-1表达及共表达情况分低、中、高三组。所有病理科诊断均参照WHO胃癌病理学及遗传学分类,由三名病理科医师共同诊断。所有患者材料均经家属同意,并有知情同意书。获道德及伦理委员会批准。
1.2 实验方法 标本经甲醛固定,常规程序化脱水、石蜡包埋、连续切片(4 μm)后,进行苏木精-伊红(HE)染色和免疫组化染色。Nrf2应用兔抗人 Nrf2多克隆抗体(Abcam公司;ab53019,稀释度1∶500)进行检测。IGF-1应用鼠抗人IGF-1单克隆抗体 (Santa Cruz公司;sc-74116,稀释度1∶400)进行检测。切片脱蜡,3%过氧化氢溶液孵育10 min除去内源性过氧化物酶。后进行抗原恢复步骤。切片应用0.01 mol/L柠檬酸缓冲液(pH6.0)浸泡,微波炉加热25 min。0.01 mol/L磷酸盐缓冲盐水(PBS;pH7.4)冲洗,常温下湿盒内正常血清覆盖切片30 min。0.01 mol/L PBS冲洗掉多余血清,常温下湿盒内应用一抗孵育45 min。然后,PBS冲洗,37℃下湿盒内应用生物酰化的二抗孵育40 min,PBS冲洗,加入链亲和素-过氧化物酶复合物(DAKO),切片在室温下孵育45 min,0.01 mol/L PBS冲洗,显微镜下3,3'二氨基联苯胺4-盐酸溶液(DAB)覆盖15 min(显色),流动的自来水浸泡,HE复染1 min,自来水浴,一系列渐高浓度酒精浸泡,而后二甲苯浸泡,封盖玻片。用PBS代替一抗作为阴性对照。
1.3 结果判断 细胞染色阳性信号为棕黄色颗粒,定位于细胞质或细胞核。组织学及免疫组化染色评价由3位病理学家独立完成。有争议的切片重新评估以达成共识。每份样本,至少3 000个细胞进行免疫组化染色评估。切片放大200倍观察,测定胞质染色的原发性癌细胞百分比。另外,根据肿瘤细胞染色的百分数,分为“低表达”(<33%癌细胞染色)、“中表达”(≥33%并且<66%癌细胞染色)及“高表达”(≥66% 癌细胞染色)〔5〕。
1.4 统计学方法 应用SPSS11.0统计软件进行分析,采用方差齐性检验,两组间比较应用Pearson相关分析及Mann-Whitney检验。
2 结果
2.1 Nrf2因子及IGF-1在非肿瘤性病变中的表达 Nrf2在正常胃黏膜几乎无表达,良性增生性病变中低表达;阳性表达多位于细胞质,部分位于细胞核;30例增生性息肉中,16例无表达,14例合并严重炎症者呈低表达。与Nrf2表达情况类似,IGF-1在正常胃黏膜组织中无表达;在增生性息肉中,2例中度表达,13例低表达,15例无表达。Nrf2及IGF-1在系列切片中,2例呈现共表达。见图1。
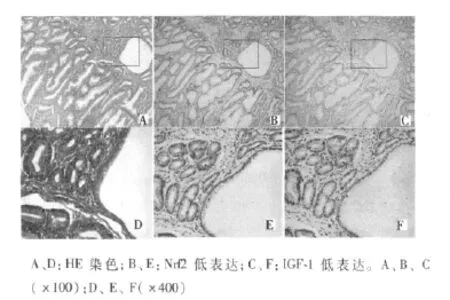
图1 增生性息肉Nrf2和IGF-1表达
2.2 Nrf2因子及IGF-1在GIN中表达 Nrf2在癌前病变中表达增强。Nrf2在62例低级别GIN中,13例高表达,25例中等表达,24例低表达(包括4例阴性);34例高级别GIN者,15例高表达,15例中表达,4例低表达(包括2例阴性);Nrf2在高级别及低级别GIN中表达差别有统计学意义(P=0.034)。IGF-1在GIN中表达上调,62例低级别GIN中,18例高表达,15例中等表达,29例低表达(包括4例阴性);34例高级别GIN者,7例高表达,6例中表达,21例低表达(包括13例阴性表达);IGF1在高级别及低级别GIN中表达差别无统计学意义(P=0.058)。Nrf2及IGF-1共表达,在GIN中表达水平较增生性息肉提高,而高、低级别GIN中高、中、低共表达分别为7、6、4例及12、21、12例,组间无明显差别(P=0.062)。见图2。
2.3 Nfr2及IGF-1在腺癌中的表达 在135例腺癌中,Nrf2、IGF-1表达均增强,Nrf2及IGF-1共表达亦增强,见图3。Nfr2、IGF-1以及共表达情况在增生性息肉、GIN及腺癌中差异有统计学意义(均P<0.05)。此外,良性病变、癌前病变及恶性病变中,Nfr2及 IGF-1表达呈正相关(r=0.337,P=0.037)。Nrf2、IGF-1及二者共表达与肿瘤分类、侵犯深度及转移有关。见表1。
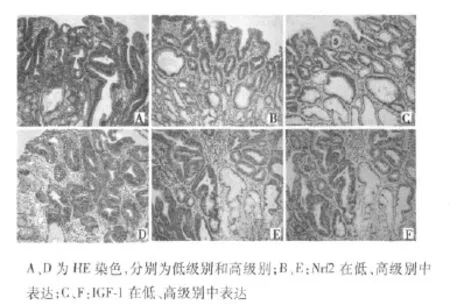
图2 上皮内肿瘤Nrf2和IGF-1表达(×200)
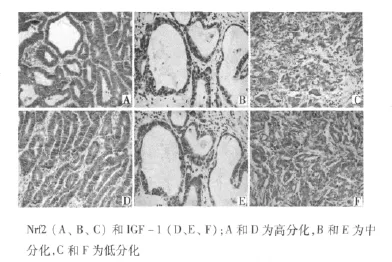
图3 腺癌Nrf2和IGF-1表达(×200)
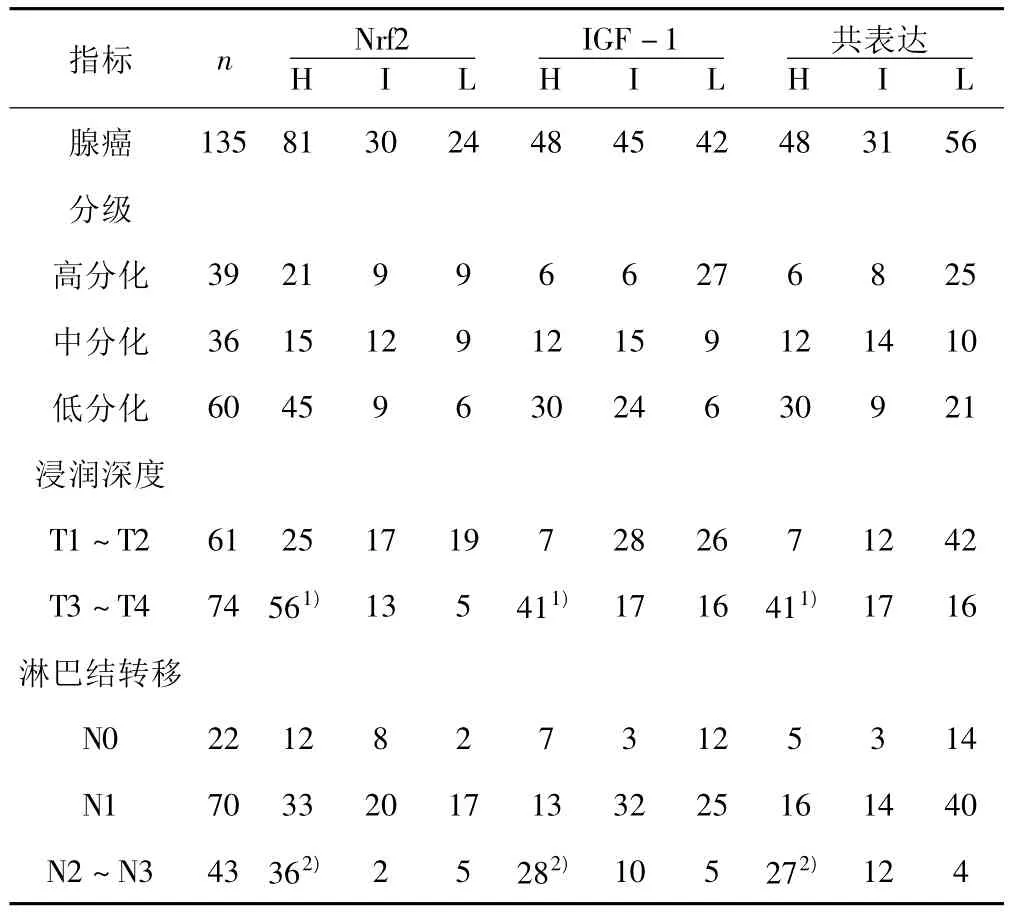
表1 Nrf2及IGF-1表达与胃腺癌浸润深度及淋巴结转移的关系(n)
3 讨论
癌变过程是环境和遗传因素相互作用而引起多基因突变和其他生物大分子改变积累的结果,是涉及启动、促进和进展等多阶段的复杂过程。化学致癌物经肝脏I相代谢酶(如细胞色素P450)催化后形成亲电子活性致癌物,与DNA及蛋白质结合形成加成物,引起癌基因和抑癌基因改变;而经Ⅱ相代谢酶,如谷胱甘肽s-转移酶(GST)的生物转化作用,使终致癌物亲水性增强,促进其体外排泄。Nrf2是调节Ⅱ相代谢酶表达的重要转录因子,通过与靶基因的抗氧化反应元件(ARE)结合,调节多种解毒酶的表达。Nrf2的激动剂CDDO-Im可以预防黄曲霉素介导的肝细胞癌前病变的发生〔6,7〕。抑制Nrf2向细胞核易位会加重氧化应激,导致非诺贝特介导的肝癌的形成〔8〕。Nrf2缺陷鼠比野生鼠更易形成肺转移癌结节〔9〕。
氧化应激破坏了机体内氧化和抗氧化的平衡,致使组织细胞中自由基产生增多,从而导致了包括肿瘤在内的一些疾病的发生。生理状态下,Nrf2与胞浆蛋白伴侣分子Keap1(Nrf2的Cullin3型泛素连接酶)耦连,锚合于胞质,处于活性相对抑制状态。亲电子脂质过氧化物4-羟基-2-壬烯醛(HNE)通过修饰Keap1,使其构象发生改变,使Nrf2解离后转入胞核〔10〕。在癌症化学预防研究中,叔丁基对苯二酚(tBHQ)及莱菔硫烷(SFN)诱导Nrf2在结直肠癌细胞胞核内积聚,激活Nrf2依赖的ARE介导的基因表达〔11〕。P62可与Keap1的Nrf2结合位点相互作用,P62过表达或自我吞噬缺陷与Nrf2和Keap1的交互作用竞争,最终导致Nrf2稳定及其靶基因转录活化〔12〕。近期一项研究表明,1 145例癌组织应用单链构象多态性分析(SSCP),其中22例鳞癌中有21例检测到Nrf2基因突变(突变率99.5%)。所有发现Nrf2突变的食管鳞癌及皮肤鳞癌中,细胞核中的Nrf2表达均增强。然而,所有鳞癌中未发现Keap1突变。以上提示Nrf2突变在鳞癌发展中具有至关重要的作用,这是鳞癌的特点〔13〕。在人类的结肠癌细胞中,一氧化氮促进Nrf2在胞核中快速积聚,对照组细胞中的Nrf2则在胞质中表达。Nrf2在胞核中积聚与转录增强和转录后修饰有关〔14〕。IGF-1是有丝分裂潜在刺激物,能促进细胞分化及转化。此外,IGF-1是主要的生存因子,可以保护细胞免于各种环境下的凋亡。
本研究证实,Nrf2和IGF-1在胃黏膜癌前病变及腺癌中表达较良性增生性病变明显升高,与此结果一致的是,二者共表达水平在胃黏膜癌前病变及腺癌中亦升高。提示二者与癌变过程甚至与肿瘤侵袭及转移有关。在正常及良性病变,二者共表达为无或低水平,但在癌前病变及腺癌中共表达上调。这一结果揭示二者之间存在协同效应,这种协同可能导致癌变发生,甚至促进肿瘤侵袭及转移。Nrf2表达水平在高、低级别GIN差异显著,提示高级别GIN可能接受了更多的氧化损伤,导致细胞DNA损伤及突变,即Nrf2基因的突变。有研究报导Nrf2为一种核蛋白〔15〕,本研究结果表明其在癌前病变及腺癌细胞质中表达,显示胞质Nrf2可能为突变后过表达并有促进癌变发生的作用。另外,IGF-1表达从良性病变至恶性病变增加,提示其有保护癌细胞抗凋亡及促进肿瘤进展作用。国外研究已证实,IGF-1阳性结直肠癌较阴性者有更高的进展率,且有较高妥昔单抗(表皮生长因子单抗)耐药率,生存期亦缩短〔4〕。本研究资料显示低分化腺癌较高分化者有更高的Nrf2和IGF-1阳性表达率。Nrf2和IGF-1共表达暗示其在胃癌发生进展方面具有协同作用,可能对预后预测有一定价值。
1 Jeong WS,Jun M,Kong AN.Nrf2:a potential molecular target for cancer chemoprevention by natural compounds〔J〕.Antioxid Redox Signal,2006;8(1-2):99-106.
2 叶社房,侯振清,钟李明,等.Nrf2/ARE在化学防癌机制中的作用〔J〕. 中华肿瘤防治杂志,2007;14(3):222-5.
3 Alberobello AT,D'Esposito V,Marasco D,et al.Selective disruption of insulin-like growth factor-1(IGF-1)signaling via phosphoinositide-dependent kinase-1 prevents the protective effect of IGF-1 on human cancer cell death〔J〕.J Biol Chem,2010;285(9) :6563-72.
4 Scartozzi M,Mandolesi A,Giampieri R.Insulin-like growth factor 1 expression correlates with clinical outcome in K-RAS wild type colorectal cancer patients treated with cetuximab and irinotecan〔J〕.Int J Cancer,2010;127(8):1941-7.
5 Dango S,Sienel W,Schreiber M,et al.Elevated expression of carcinoembryonic antigen-related cell adhesion molecule 1(CEACAM-1)is associated with increased angiogenic potential in non-small-cell lung cancer〔J〕.Lung Cancer,2008;60(3):426-33.
6 Roebuck BD,Curphey TJ,Li Y,et al.Evaluation of the cancer chemopreventive potency of dithiolethione analogs of oltipraz〔J〕.Carcinogenesis,2003;24(12):1919-28.
7 Yates MS,Kwak MK,Egner PA,et al.Potent protection against aflatoxininduced tumorigenesis through induction of Nrf2-regulated pathways by the triterpenoid 1-〔2-cyano-3-,12-dioxooleana-1,9(11)-dien-28-oyl〕imidazole〔J〕.Cancer Res,2006;66(4):2488-94.
8 Nishimura J,Dewa Y,Okamura T,et al.Role of Nrf2 and oxidative stress on fenofibrate-induced hepatocarcinogenesis in rats〔J〕.Toxicol Sci,2008;106(2):339-49.
9 Satoh H,Moriguchi T,Taguchi K,et al.Nrf2-deficiency creates a responsive microenvironment for metastasis to the lung〔J〕.Carcinogenesis,2010;31(10):1833-43.
10 Mahaffey CM,Zhang H,Rinna A,et al.Multidrug-resistant protein-3 gene regulation by the transcription factor Nrf2 in human bronchial epithelial and non-small-cell lung carcinoma〔J〕.Free Radic Biol Med,2009;46(12):1650-7.
11 Wu XY,Qu LY,Quan K,et al.Effect of tBHQ and sulforaphane on Nrf2-ARE signaling pathway of cancer cells〔J〕.Zhejiang Da Xue Xue Bao Yi Xue Ban,2010;39(1):17-23.
12 Komatsu M,Kurokawa H,Waguri S,et al.The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1〔J〕.Nat Cell Biol,2010;12(3):213-23.
13 Kim YR,Oh JE,Kim MS,et al.Oncogenic NRF2 mutations in squamous cell carcinomas of oesophagus and skin〔J〕.J Pathol,2010;220(4):446-51.
14 Li CQ,Kim MY,Godoy LC,et al.Nitric oxide activation of Keap1/Nrf2 signaling in human colon carcinoma cells〔J〕.Proc Natl Acad Sci U S A,2009;106(34):14547-51.
15 Nguyen T,Sherratt PJ,Nioi P,et al.Nrf2 Controls Constitutive and in-ducible expression of ARE-driven genes through a dynamic pathway involving nucleocytoplasmic shuttling by Keap1〔J〕.J Biol Chem,2005;280(37):32485-92.
R735.2
A
1005-9202(2012)17-3658-04;
10.3969/j.issn.1005-9202.2012.17.018
1 山东大学第二医院消化科
王洪波(1974-),男,博士,主治医师,主要从事消化道肿瘤基础及临床研究。
宋素贞(1977-),女,硕士,讲师,主要从事消化道肿瘤基础及临床研究。
〔2012-01-07收稿 2012-03-04修回〕
(编辑 袁左鸣)
猜你喜欢
安徽农业科学(2022年1期)2022-02-14
江苏农业学报(2021年2期)2021-06-30
癌变·畸变·突变(2021年2期)2021-04-15
昆明医科大学学报(2020年12期)2021-01-26
癌变·畸变·突变(2020年5期)2020-10-19
科学与财富(2020年15期)2020-07-04
移动通信(2020年4期)2020-05-07
中国临床医学影像杂志(2019年6期)2019-08-27
移动通信(2019年4期)2019-06-25
现代信息科技(2018年4期)2018-07-12
