
固氮基因的分子进化分析
2013-11-05 06:40陈海魁王俊丽
江西农业大学学报 2013年3期
王 前,陈海魁,王俊丽
(1.中央民族大学 生命与环境科学学院,北京100081;2.北方民族大学 生物科学与工程学院,宁夏 银川750021;3.北京市八达岭林场,北京102112)
经典分类学主要通过研究化石证据、形态解剖学和生理学等方面来建立生物进化的总体模式以及勾画物种之间的谱系关系。自20 世纪中后期,计算机科学和生物技术取得了迅猛发展,分子遗传学资料迅速积累,使得分子进化逐渐被应用于生物进化研究[1-2]。分子进化是从分子水平上研究进化,通过对核酸和蛋白质大分子序列分析,能够解析出其中蕴含的进化信息,推断其进化历史,为生物进化提供重要的依据[3]。
生物固氮是氮元素进入生态系统物质与能量循环的重要途径,根据固氮微生物与其它生物的关系,可将生物固氮分为自生固氮、联合固氮和共生固氮3 种不同的体系[4-5],其中根瘤菌与豆科植物的共生固氮是自然界中主要的固氮体系[6],在固氮微生物的固氮基因中,以根瘤菌为例,参与根瘤发育的固氮基因分为3 类:第1 类是结瘤基因,如nol 基因、nod 基因和noe 基因;第2 类是与根瘤菌细胞表面结构有关的基因,如脂多糖基因(lps 基因)、胞外多糖基因(exo 基因)、和ndv 基因;第3 类是固氮酶基因(简称nif 基因)和共生固氮基因(fix 基因)[7-9]。本文研究了根瘤菌和豆科植物共生作用模式菌苜蓿中华根瘤菌(Sinorhizobium meliloti 1021)中的3 类固氮基因的进化关系。
nifH 基因负责编码固氮酶组分铁蛋白亚基,为固氮微生物所共有,具有高度保守性,是科学家研究生物固氮的重点领域之一[10]。在过去几年中,越来越多固氮基因(nif 基因)已被确定,Renato Fani 等通过研究nifD,nifK,nifE,nifN,nifH 基因的进化历史,了解固氮基因的分子进化[11]。张于光等[12]利用PCR-RFLP 和测序分析对青海三江源高寒草甸土壤微生物固氮基因(nifH)的多样性和系统发育进行了研究。马驿等[13]通过PCR 扩增、基因克隆等方法对恩诺沙星影响下的土壤微生物固氮酶nifH 基因进行了分子多样性分析。
在根瘤菌与豆科植物形成的共生固氮体系中,根瘤菌与豆科植物通过协同作用固氮。豆科植物的固氮基因enod40 为早期结瘤素基因(简称enod)的一种,参与根瘤形成与发育,它与其他固氮基因一起参与协同固氮[14-15]。据报道,enod40 基因在根瘤菌器官形成过程中起着重要的作用,在多种植物中该基因编码的核酸序列有较高的保守性[16-18]。
目前,国内外已有不少关于固氮基因分离、鉴定、定位、克隆、表达调控机理等方面的研究[19-21],但在固氮基因进化方面的研究较少[22]。本文从同一固氮微生物固氮基因、同一固氮基因在不同固氮微生物和不同固氮植物3 个方面进行固氮基因的分子进化分析,在分子水平上研究了生物间的亲缘关系,并绘制出生物进化分子树,为进一步深入研究固氮基因的进化机制提供了重要依据。
1 材料与方法
1.1 供试材料
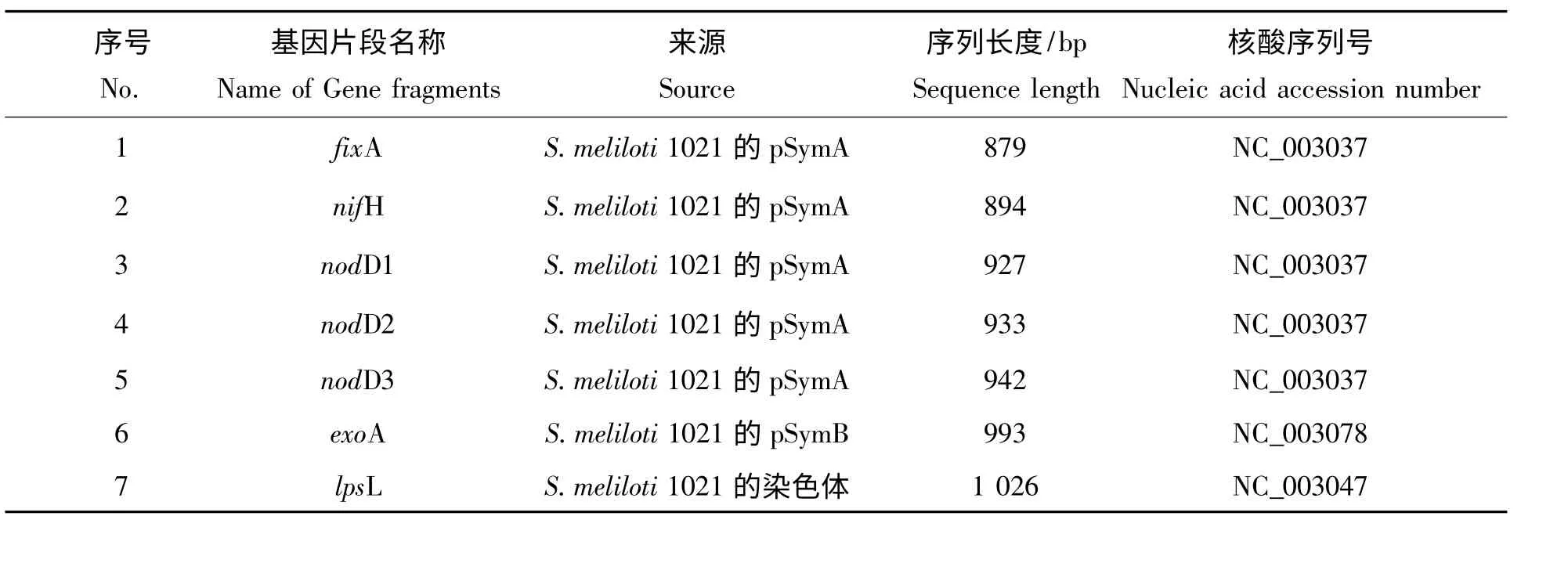
表1 S.meliloti 1021 需要作分子进化分析的基因片段Tab.1 S.meliloti 1021 gene fragments used in the molecular evolution analysis
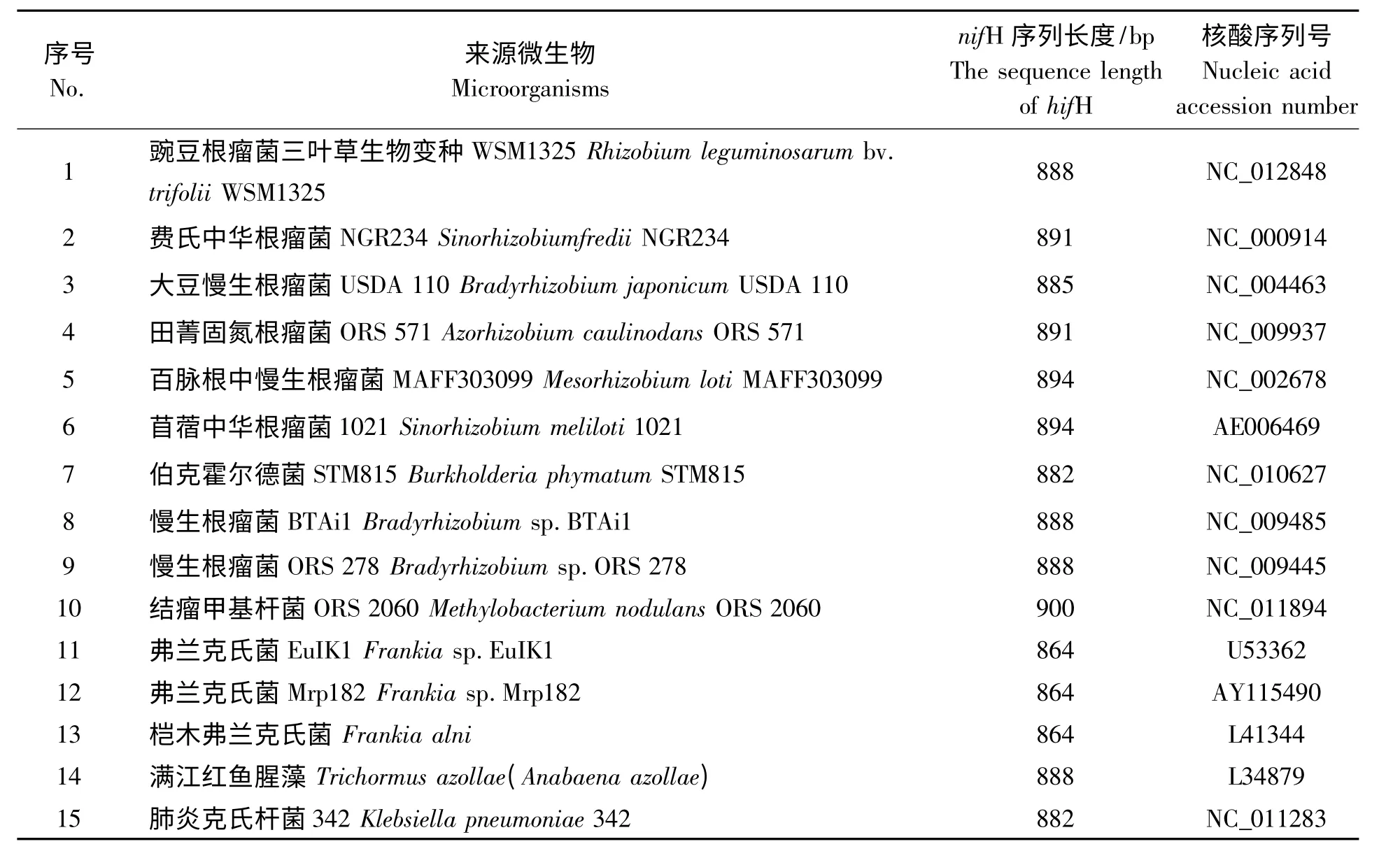
表2 不同固氮微生物nifH 基因序列Tab.2 The different nifH gene fragments of nitrogen-fixing microorganisms
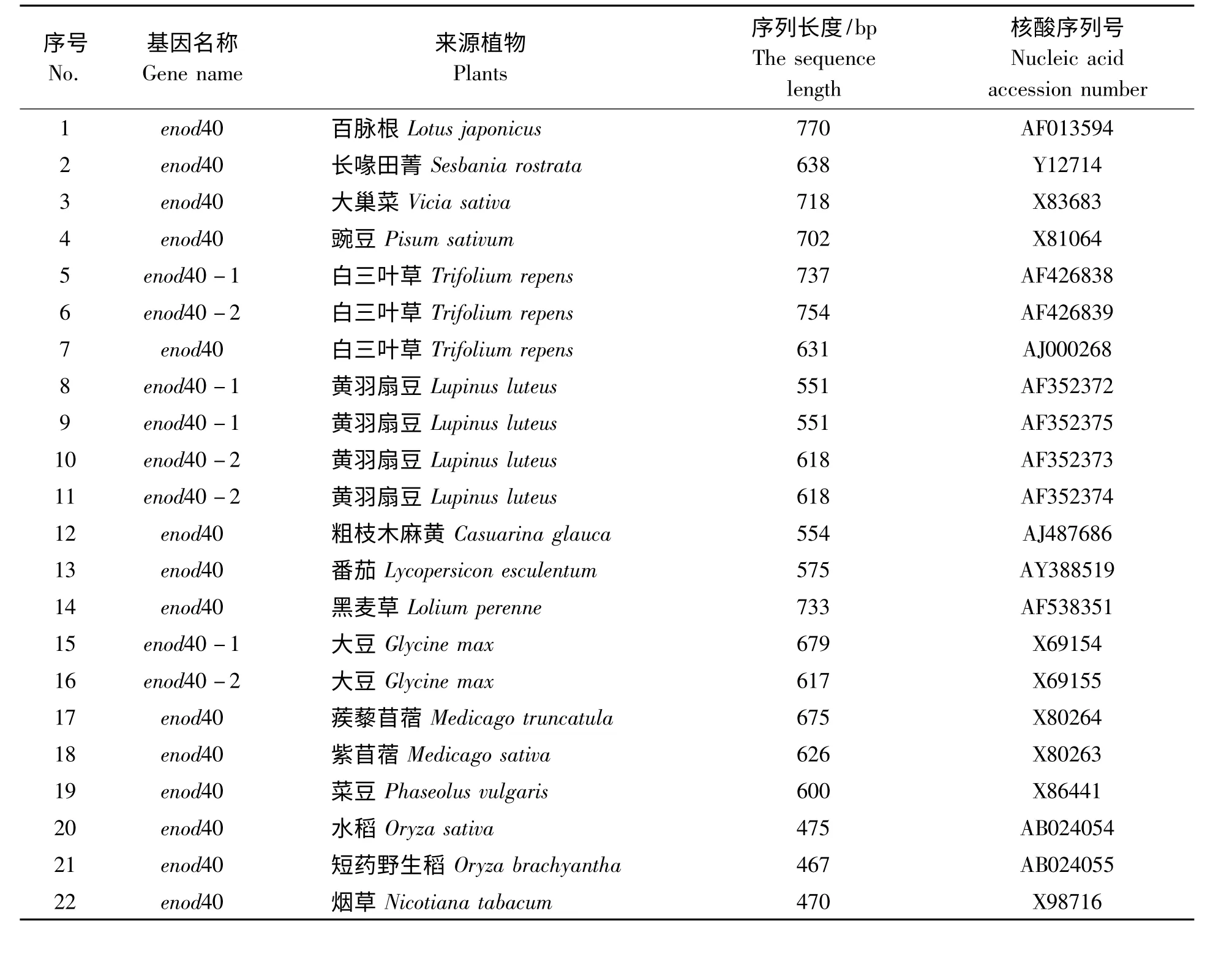
表3 不同固氮植物enod40 基因序列Tab.3 The different enod40 gene fragments of nitrogen-fixing plants
1.1.1 Sinorhizobium meliloti 1021 固氮基因序列的获得与整理 登录美国国立生物技术信息中心(NCBI)网站主页,在GenBank 中搜寻Sinorhizobium meliloti 1021 固氮基因序列,最终用作分子进化分析的固氮基因序列(表1)。
1.1.2 固氮微生物nifH 基因的获得与整理 登录美国国立生物技术信息中心(NCBI)网站主页,在GenBank 中搜寻豌豆根瘤菌、费氏中华根瘤菌、大豆慢生根瘤菌、田菁固氮根瘤菌等固氮微生物nifH 基因序列,最终用作分子进化分析的固氮基因序列(表2)。
1.1.3 固氮植物enod40 基因的获得与整理 登录美国国立生物技术信息中心(NCBI)网站主页,在GenBank 中搜寻百脉根、大豆、大巢菜、豌豆等固氮植物enod40 基因序列,最终用作分子进化分析的固氮基因序列(表3)。
1.2 方法
先用BioEdit7.0 从核酸序列中找到需要作分子进化分析的固氮基因部分并分别保存到3 个文件中;利用Clustal X1.83 对序列进行完全比对;再用BioEdit7.0 将输出的比对文件(aln 格式)转换成fas 格式文件;最后导入MEGA4.0 软件,通过邻接法(Neighbor -Joining,简称NJ),经过自展法(Bootsrap)1 000 次重复验证,以保证树拓扑结构的可靠性,构建基因分子进化树。
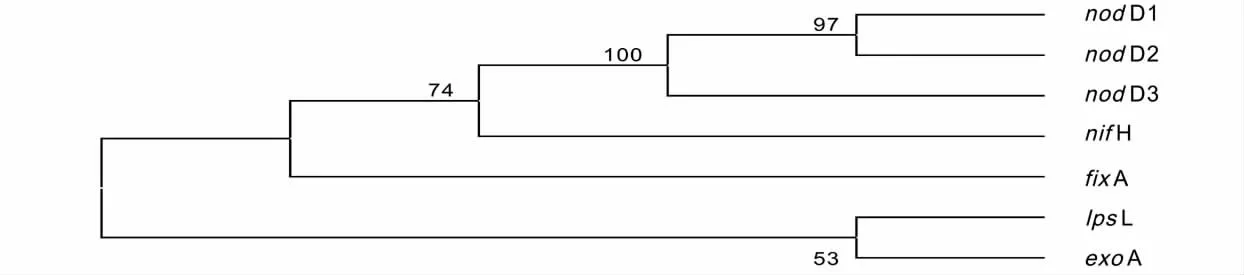
图1 Sinorhizobium meliloti 1021 固氮基因进化树Fig.1 Phylogenetic tree of nitrogen-fixing gene in Sinorhizobium meliloti 1021
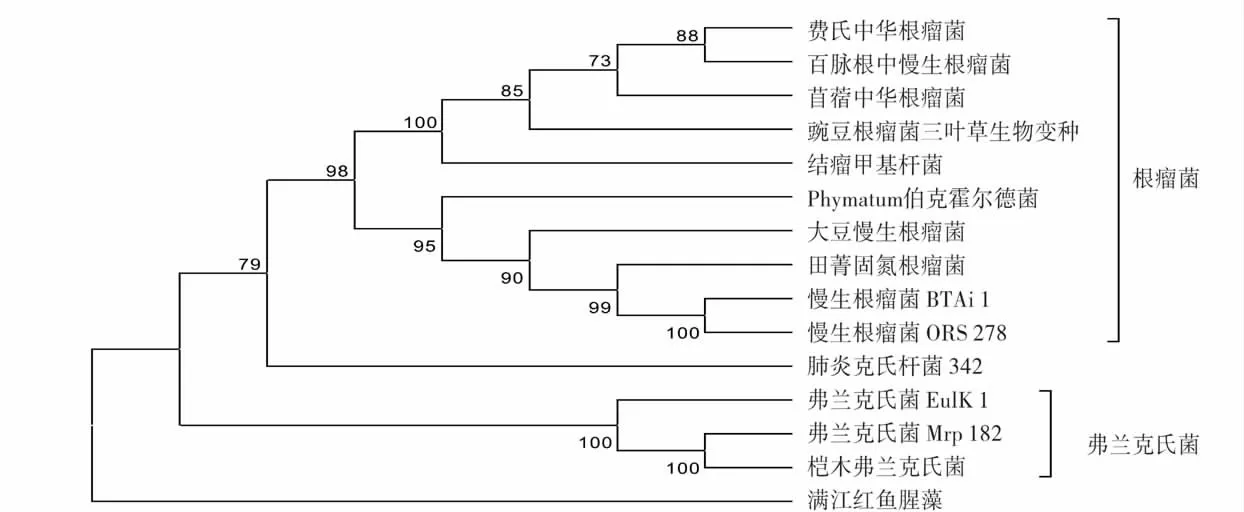
图2 固氮微生物nifH 基因进化树Fig.2 Phylogenetic tree of nifH gene in nitrogen-fixing microorganisms
2 结果与分析
2.1 Sinorhizobium meliloti 1021 固氮基因进化树分析
从Sinorhizobium meliloti 1021 固氮基因进化树(图1)可知,除了lpsL 和exoA 基因间的自展值(bootstrap 值)小于70%外,其它基因间均大于70%,因此该进化树可认为是可靠的。不同结瘤基因间的差异极小,自展值达100%。固氮基因可分为固氮酶基因(nif 基因)和共生固氮基因(fix 基因),由于功能的不同而分为两枝,nifH 与nod 基因间的自展值为74%。exoA 和lpsL 均为根瘤菌细胞表面结构有关的基因,由于exoA 为糖基转移酶基因,lpsL 为脂多糖基因,功能的不同造成两者差异较大,自展值为53%。该进化树分枝与根瘤菌中参与共生固氮的基因分类分为结瘤基因(nod、nol、noe 基因等)、与根瘤菌细胞表面结构有关的基因(如exo、lps、ndv 基因等)和固氮酶基因3 类相符合,因为结瘤与固氮在生物固氮过程中密切相关,在实际的根瘤菌固氮基因的进化中,能否认为nod、nif 和fix 基因为一个分枝,lps 和exo基因为一个分枝,还有待于进一步研究。

图3 固氮植物enod40 基因进化树Fig.3 Phylogenetic tree of enod40 gene in nitrogen-fixing plants
2.2 固氮微生物nifH 基因进化树分析
从固氮微生物固氮基因(nifH)进化树(图2)可以看到,自展值为73% ~100%,大于70%,因此该进化树是可靠的。该进化树分为4 枝,根瘤菌为一枝,nifH 进化树中不同根瘤菌的分枝与目前根瘤菌16S rRNA 系统发育树的分类相比较还有一定差别[23],伯克霍尔德氏菌和根瘤菌聚合成为一枝,研究表明伯克霍尔德氏菌具有固氮的能力[24],但其固氮方式未见报道,分析发现伯克霍尔德氏菌与根瘤菌的nifH 基因序列相似度较大,其固氮方式是否为共生固氮还有待进一步研究;肺炎克氏杆菌为一枝,它是兼性厌氧自身固氮微生物;弗兰克氏菌为一枝,其nifH 基因进化较为一致,自展值为100%;满江红鱼腥藻为一枝,这可能是它在固氮方式上与根瘤菌和弗兰克氏菌有很大差异。
2.3 固氮植物enod40 基因进化树分析
从固氮植物固氮基因(enod40)进化树(图3,以单子叶植物的禾本科为外类群)看出,自展值基本上大于70%,认为该进化树是可靠的。该进化树分为4 枝,大豆、菜豆、长喙田菁百脉根等10 种植物聚为一枝,自展值为89%;番茄和烟草聚为一枝,自展值为100%;黑麦草、水稻和短药野生稻聚为一枝,自展值为100%;粗枝麻黄独自为一枝。从另一方面看,这4 分枝刚好为4 个科,与APGIII 的分类基本吻合[25]。通过分析得出,enod40 基因进化树将植物分为双子叶植物和单子叶植物两大类,双子叶植物主要通过与固氮微生物共生固氮,而禾本科植物通过联合固氮利用大气中的氮元素[26],它们都共同有植物早期结瘤素基(enod40),表明enod40 基因为较为原始的基因,在固氮过程中起着重要作用。
3 讨论
本研究首次以根瘤菌和豆科植物共生作用模式菌苜蓿中华根瘤菌(Sinorhizobium meliloti 1021)里的不同固氮基因序列以及从GenBank 数据库中检索出的其他固氮微生物的nifH 基因和不同植物中的enod40 基因序列为研究对象,通过邻接法构建系统进化树,从分子水平上确定了固氮基因、固氮微生物、固氮植物之间亲缘关系和生物进化顺序,为固氮生物进化论的研究提供了更为客观、精确、定量的研究结论,促进了固氮基因进化方面的研究发展。
通过固氮微生物nifH 基因进化树和固氮植物enod40 基因进化树分析,得出的分类结果和目前的分类系统较为一致,可以认为,用邻接法分析保守固氮基因序列构建的分子进化树是有意义的,本研究有助于了解各类固氮生物之间的进化关系和不同生物的固氮机制研究。
[1]向福.红豆杉的18S rRNA 基因的分子进化研究[D].武汉:华中科技大学,2004.
[2]阎锡海,王延峰.分子树在进化论研究中的作用[J].青海师范大学学报:自然科学版,1999,20(1):48 -52.[3]王禄山,高培基.生物信息学应用技术[M].北京:化学工业出版社,2008.
[4]黄群策,陈启锋,李志真.生物固氮研究的前景[J].科技导报,1999,19(1):26 -29.[5]陈今朝,向邓云.生物固氮的研究与应用[J].涪陵师专学报,2000,16(2):93 -95.
[6]韩斌,孔继君,邹晓明,等.生物固氮研究现状及展望[J].山西农业科学,2009,37(10):86 -89.[7]吕成群.相思树根瘤菌的研究[D].南京:南京林业大学,2004.
[8]冯春生.西北地区天蓝苜蓿根瘤菌系统发育研究[D].咸阳:西北农林科技大学,2008.
[9]周俊初.根瘤菌共生固氮基因的结构、功能和调控[J].高技术通讯.1996,11(7):59 -62.
[10]文都日乐,李刚,杨殿林,等.呼伦贝尔草原土壤固氮微生物nifH 基因多样性与群落结构[J].生态学杂志,2010,30(4):790 -797.
[11]Renato Fani,Romina Gallo,Pietro Lio.Molecular evolution of nitrogen fixation:The evolutionary history of the nifD,nifK,nifE,and nifN Genes[J].Journal of Molecular Evolution,2000,51:1 -11.
[12]张于光,王慧敏,李迪强,等.三江源高寒草甸土固氮基因(nifH)的多样性和系统发育研究[J].微生物学报,2005,45(2):166 -171.
[13]马驿,陈杖榴.恩诺沙星残留对土壤中固氮细菌固氮基因(nifH)多样性的影响[J].生态学报,2010,30(1):7 -14.
[14]窦新田.豆科植物—根瘤菌共生固氮体系中的根瘤菌结瘤基因组(nod 和hsn 基因)和固氮基因组(nif 和fix 基因)及其功能[J].生物技术,1992,2(3):1 -3.
[15]陈大松.紫云英共生固氮相关基因在铵胁迫及盐胁迫下的表达研究[D].武汉:华中农业大学,2007.
[16]陈卫民.西北干旱半干旱地区骆驼刺、黄芪及外来物种-刺槐根瘤菌多样性和系统发育研究[D].咸阳:西北农林科技大学,2009.
[17]万曦,Vleghels I,Franssen H,et al. 克隆植物早期结瘤素基因enod40 的受体基因[J]. 农业生物技术学报,2001,9(3):293 -296.
[18]王彦章,俞冠翘,朱家壁.水稻中结瘤素基因的同源基因研究[J].中国水稻科学,2005,19(3):202 -208.
[19]Reddy P M,Aggarwal R K,Ramos M C,et al.Widespread occurrence of the homologue of the early nodulin (enod)genes in Oryza species and related grasses[J].Biochem Biophys Res Comm,1999,258:148 -154.
[20]张执欣.西北干旱半干旱地区甘草根瘤菌遗传多样性研究[D].咸阳:西北农林科技大学,2006.
[21]何新华,陈力耕,陈怡.非豆科木本植物与放线菌共生固氮相关基因研究进展[J]. 浙江林学院学报,2004,21(1):110 -114.
[22]沈世华,荆玉祥.中国生物固氮研究现状和展望[J].科学通报,2003,48(6):535 -540.
[23]汪恩涛,曲春枫,冯继东,等.16S rRNA 基因测序确定根瘤菌的系统发育[J].高技术通讯,1995,4(11):55 -57.
[24]赵乃昕,彭效祥,高淑萍,等.椰毒伯克霍尔德氏菌-新发现的固氮菌及其简易鉴定方法[J].潍坊医学院学报,2003,25(2):93 -95.
[25]APG(The Angiosperm Phylogeny Group). An update of the angiosperm phylogeny group classification for the orders and families of flowering plants:APGIII[J].Bot J Linn Soe,2009,161:105 -121.
[26]张丽梅,方萍,朱日清.禾本科植物联合固氮研究及其应用现状展望[J].应用生态学报,2004,15(9):1650 -1653.
猜你喜欢
生物技术通报(2022年3期)2022-05-10
农业科技与信息(2021年16期)2021-12-07
生态科学(2021年5期)2021-11-13
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
园艺与种苗(2021年1期)2021-03-16
科学与技术(2019年21期)2019-04-27
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
植物营养与肥料学报(2012年5期)2012-10-26
