
广西百色烟区烟草青枯病菌的遗传多样性分析
2017-05-30 02:29史国英曾泉周文亮赖洪敏农泽梅岑贞陆胡春锦
南方农业学报 2017年5期
史国英 曾泉 周文亮 赖洪敏 农泽梅 岑贞陆 胡春锦


摘要:【目的】分析廣西百色烟区烟草青枯病菌的遗传多样性,为了解该烟区青枯病的发生流行规律和该病原菌的致病机制提供参考。【方法】通过致病力测定、生化型鉴定及BOX-PCR对69株来源于广西烟草主产区百色烟区的青枯菌株的遗传多样性进行系统分析。【结果】供试菌株存在明显的致病力分化,强致病力菌株、中等致病力菌株和弱致病力菌株出现频率分别为17.39%、62.32%和20.29%,其中以中等致病力菌群为优势菌群;供试菌株的生化型复杂,其中37株属于生化型III或生化亚型III-1、III-3和III-4,23株属于生化型Ⅰ,1株属于生化型Ⅱ,另有8株属于非标准生化型;BOX-PCR分析结果表明百色烟草青枯病菌存在丰富的遗传多样性,聚类分析结果显示菌株的遗传多态性与菌株的地理来源、致病力、生化型具有一定的相关性,但并无明显的地理种群、生化型种群或致病力一致的种群聚在一起。【结论】广西百色烟区烟草青枯病菌具有丰富的遗传多样性和复杂的生化型,可能是该烟区烟草青枯病为害逐年加重的重要原因之一。
关键词: 烟草;青枯菌;生化型;遗传多样性;广西百色
中图分类号: S432.4 文献标志码:A 文章编号:2095-1191(2017)05-0825-06
Genetic diversity of Ralstonia solanacearum isolated
from tobacco in Baise, Guangxi
SHI Guo-ying 1, ZENG Quan1, ZHOU Wen-liang 2, LAI Hong-min2, NONG Ze-mei1,3,
CEN Zhen-lu1 *, HU Chun-jin1 *
(1 Microorganism Research Institute, Guangxi Academy of Agricultural Sciences, Nanning 530007, China;
2 Baise Branch, Guangxi Tobacco Corporation,Baise, Guangxi 533000, China; 3 College of Agriculture,
Guangxi University, Nanning 530004, China)
Abstract:【Objective】Genetic diversity analysis for Ralstonia solanacearum isolated from tobacco in Baise, Guangxi was studied in order to explore occurrence and epidemic regulation in Baise tobacco planting region and pathogenic mechanism of R. solanacearum. 【Method】Sixty-nine representative strains of R. solanacearum isolated from different places in Baise tobacco planting area of Guangxi were analyzed for genetic diversity by pathogenicity and biovar determination, and Box-PCR analysis. 【Result】All the tested strains presented obvious pathogenicity differentiation. The proportions belonging to high, moderate and low pathogenicity were 17.39%, 62.32% and 20.29% respectively. Strains with moderate pathogenicity were dominant ones. Base on biovar test, the biovar of the tested strains were complex. Thirty-seven strains belonged to biotype Ⅲ or subtype biochemical Ⅲ-1, Ⅲ-3 and Ⅲ-4; twenty-three were biotype Ⅰ; one belonged to biotype Ⅱ and eight belong to atypical biovar. BOX-PCR analysis indicated that there was rich genomic diversity among all the tested R. solanacearum strains. According to clustering analysis, genetic diversity was related to geographical origin, pathogenicity and biovar to some extents. But there were no strain clusters with the same geographical origin, pathogenicity and biovar. 【Conclusion】R. solanacearum isolated from Baise, Guangxi has rich genetic diversity and complex biovar, and it may be one of the reasons why bacterial wilt causes more severe damages year by year in Baise tobacco planting area.
Key words: tobacco; Ralstonia solanacearum; biovar; genetic diversity; Baise, Guangxi
0 引言
【研究意义】百色是广西最大的烤烟主产区,由于受气候及长期连作等因素影响,近年来百色部分烟区不同程度地受到青枯病(Ralstonia solanacearum)为害,且常与黑胫病(Phytophthora nicotianae)、根黑腐病(Thielaviopsis basicola)等多种根茎类病害综合发生,烟农损失惨重,严重制约了广西烤烟产业的健康发展(王五权等,2015)。烟草青枯病主要发生在旱地烟,水旱轮作的烟田发病较少,迄今为止广西旱地烟的青枯病发病率近乎100%,影响了旱地烟种植面积的进一步扩大。该病是由茄科雷尔氏菌(R. solanacearum)引起的一种重要土传植物病害,寄主范围广泛(Remenant et al.,2010;蔡刘体等,2016)。由于其种群复杂,不同菌株在寄主范围、生化型、致病力等特性上存在明显差异,因而抗病或耐病品种通常随着种植年限、地域及栽培方式的改变而丧失品种抗性(Khakvar et al.,2008;Khakvar,2009;肖田等,2015;李小杰等,2016)。因此,研究烟草青枯病菌的遗传多样性对于了解该病的发生和流行规律,从而针对性地制定相应防治策略具有十分重要的意义。【前人研究進展】前人的研究表明,烟草青枯病菌存在显著的遗传多样性。郑向华等(2007)、李小杰等(2016)运用RAPD分子标记技术分别对采集自广东省韶关市和梅州市的38株青枯病菌及采集自豫南烟区的19株青枯病菌进行遗传多样性分析,结果显示这些病菌DNA具有丰富的遗传多样性,且遗传相似性与地理来源有一定的相关性。徐进等(2010)对采集自福建省南平、三明和龙岩地区的45株烟草青枯病菌进行青枯菌演化型复合PCR及对3种双糖和3种己醇氧化利用能力检测,结果显示全部菌株均为青枯菌演化型I型即亚洲分支菌株,其中43株菌株属于青枯菌生化变种III,1株菌株属于生化变种IV,1株菌株属于非标准型生化变种。Li等(2016)对我国西南、东南、长江中上游和黄淮烟区的烟草青枯病菌的遗传多样性和致病力进行研究,发现这些烟区的烟草青枯病菌均为演化型I,并可分为7个序列变种;致病力分析结果显示菌株存在高、中、低3种致病型分化,但菌株的致病力与其序列变种和演化型间没有明显相关性;研究证明我国不同地理来源的烟草青枯菌表现出不同的遗传多样性水平,地理分布越往北,烟草青枯菌的遗传多样性越低。最近,曹毅等(2017)通过高通量测序分析了烟草青枯病与土壤细菌群落结构的关系,证明烟草青枯病病圃土壤中的拟杆菌门和放线菌门细菌在总菌群中丰度较低,研究结果为有针对性地筛选青枯病高效生防资源、通过微生物区系的生态调控控制青枯病为害提供了理论依据。【本研究切入点】本课题组前期已对广西百色烟草青枯病发生为害情况进行了全面调查和病原菌鉴定,证明百色烟草青枯菌在演化型分类上均属于演化型I(王五权等,2015),但目前针对百色烟草青枯病菌遗传多样性及其生化型尚无系统地研究报道,当地烟草品种的推广和布局缺乏依据,病害防治工作极其被动。【拟解决的关键问题】在前期工作基础上,通过致病力测定、生化型鉴定及BOX-PCR分析等技术手段,对来源于广西百色烟区的69株青枯菌的遗传多样性进行系统研究,为了解该烟区青枯病的发生流行规律和该病原菌的致病机制提供参考。
1 材料与方法
1. 1 试验材料
供试烟草青枯病菌菌株共69株,均为本课题组于2014~2015年间从百色市靖西、乐业、德保和凌云县等烟草主产区的典型病株上分离获得,所有菌株均已经过分离纯化、致病性测定及青枯菌演化型分类鉴定等研究,所有供试菌株均属于演化型Ⅰ(王五权等,2015)。供试烟草品种为云烟97。
1. 2 菌株致病力测定
供试菌株用无菌水配成3×107 CFU/mL的菌悬液备用。取5~6叶期云烟97供试。参考岑贞陆等(2013)的针刺接种法接种病原菌,接种后逐日观察、测量并记录菌株接种产生病斑扩展长度,直到病情稳定不再进一步扩展为止。
根据接种点茎秆的病斑扩展长度确定菌株的相对致病力。将菌株的致病力分为强、中、弱3个评价等级:强致病力(H),病斑平均扩展长度>5 cm;中等致病力(M),5 cm≥病斑平均扩展长度≥2 cm;弱致病力(L),病斑平均扩展长度<2 cm。
1. 3 菌株生化型测定
参考亢雅娟等(2008)、徐进等(2010)的青枯菌生化型测定方法,分析所有供试菌株对3种双糖(麦芽糖、乳糖和纤维二糖)和3种己醇(山梨醇、甘露醇和甜醇)的利用能力及对硝酸盐的还原情况,根据青枯菌生化型的划分标准(表1)确定菌株的生化型。
1. 4 菌株BOX-PCR多态性分析
利用细菌基因组DNA提取试剂盒提取供试菌株的基因组DNA。BOX-PCR分析参考林海云等(2011)的方法。PCR反应体系25.0 μL:10.0 μL ddH2O;2×Taq MasterMix 13.0 μL,10 μmol/L BOXA1R引物1.0 μL;1.0 μL模板DNA(约50 ng)。扩增程序:94 ℃预变性5 min;94 ℃1 min,52 ℃ 1 min,68 ℃ 7 min,进行32个循环;最后68 ℃延伸15 min。扩增产物采用2%琼脂糖进行电泳,电泳时间1 h,用UVP凝胶成像仪拍摄。
BOX-PCR电泳图谱分析:参照BOX扩增结果的琼脂糖凝胶电泳图谱,每一条DNA带均为一个分子标记,并代表一个引物结合位点。根据各分子标记的迁移率及其有无统计所得位点的二源数据(对电泳图上同一位置无电泳条带的计为0,有条带的计为1)记录谱带。菌株遗传相似系数的计算和系统聚类分析通过DPS 7.05完成。
2 结果与分析
2. 1 供试菌株的致病力分化情况
根据菌株接种发病及病斑扩展情况和致病力评价标准,供试69株菌株在云烟97茎秆上致病性反应可分为强致病力、中等致病力和弱致病力3种类型(图1),强致病力菌株、中等致病力菌株和弱致病力菌株出现的频率分别为17.39%、62.32%和20.29%(表2),其中以中等致病力菌群为优势菌群。
2. 2 供试菌株的生化型鉴定结果
根据菌株对3种双糖、3种已醇及对硝酸盐还原的反应模式,发现69株供试菌株中有37株属于生化型Ⅲ或Ⅲ-1、Ⅲ-3、Ⅲ-4等生化亚型,23株属于生化型Ⅰ,1株属于生化型Ⅱ,另有8株无法归属于前人划分的生化型反应模式,属于非标准型生化变种(W)(表2)。
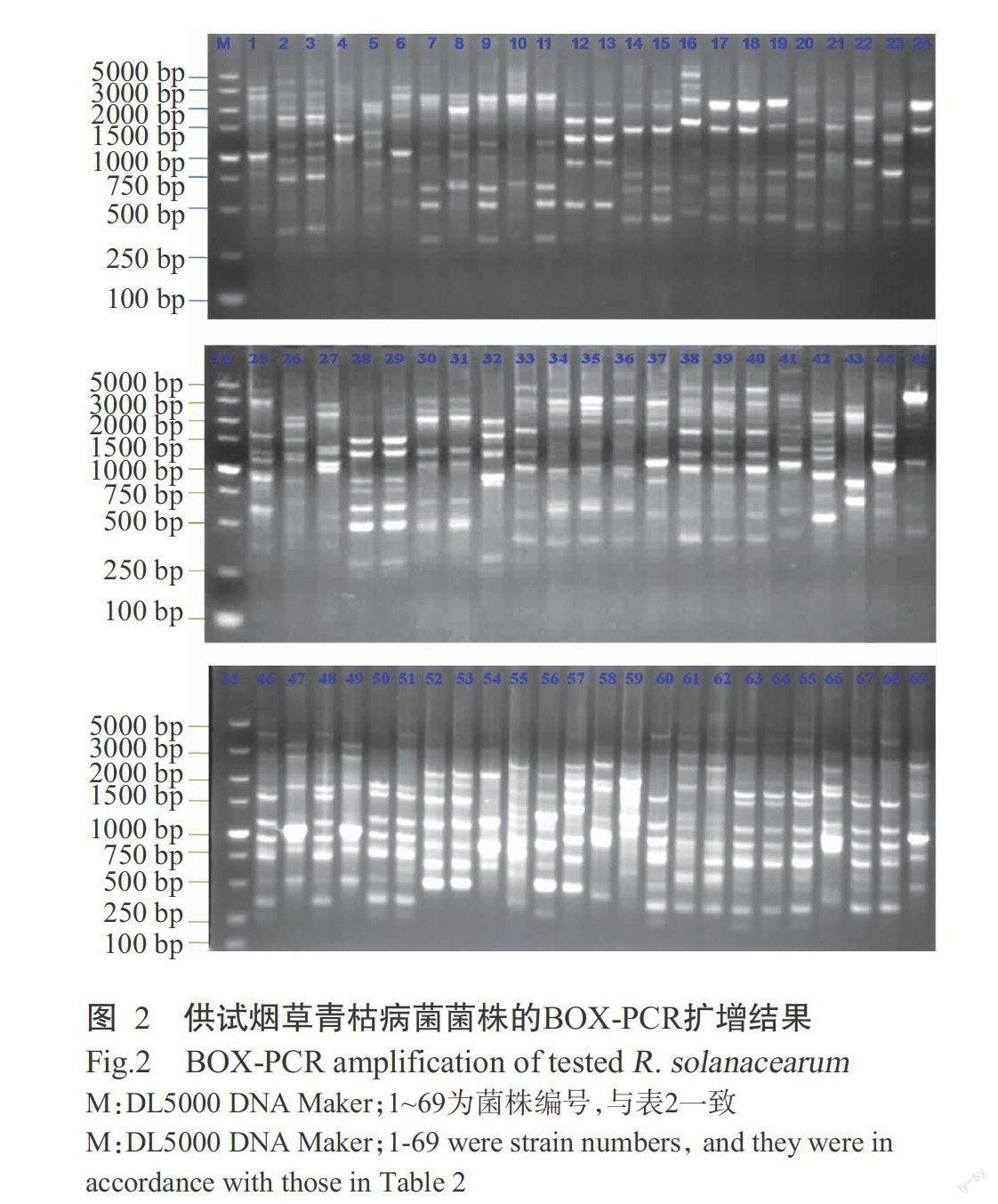
2. 3 供试菌株BOX-PCR多态性分析结果
供试烟草青枯菌菌株的BOX-PCR扩增结果电泳分析结果见图2,采用BOX引物共扩增出18种条带,且全部为特异性条带,多态带比率为100%。
对所有供试菌株的BOX-PCR扩增结果进行分析,电泳图上扩增条带的有无分别记为1和0,建立“0,1”表,将“0,1”数据输入统计分析软件DPS 7.05进行聚类分析,获得由不同菌株基因组指纹图谱构建的聚类分析树状图(图3)。聚类分析结果表明,百色烟草青枯病菌存在丰富的遗传多样性,几乎每个菌株都有各自的多样性条带,69株菌株在遗传距离为0.60的基础上聚为一群。菌株的聚类分析结果与菌株来源有一定的相关性,如少数同源性极高的菌株均来源于同一烟区,但有部分来源相同的青枯菌的遗传距离较远;同时,遗传多样性与菌株的生化型、致病力等也存在一定的相关性,如编号为12和13号菌株,17和18号菌株,46和48号菌株,67和68号菌株,它们在地理来源、生化型、致病力及多态性分析结果上均完全一致;但也有部分多态性分析结果基本一致的菌株,其生化型及致病力存在明显差异,如编号为2和3号菌株,14和15号菌株,28和29号菌株,34、35和36号菌株,52和53号菌株,63、64和65号菌株。
3 讨论
本研究供试菌株均分离自广西百色烟草主产区的青枯病植株,经过柯赫法则证明为烟草青枯病致病菌,菌株来源基本涵盖了所有发生青枯病的烟区,其结果能够反映百色烟草青枯病的遗传分化情况,对该烟区烟草青枯病菌的遗传多样性具有一定的代表性。
前人研究证明我国大部分烟区烟草青枯病菌主要含生化型III和IV(徐进等,2010;谢锐鸿等,2014),而百色烟草青枯菌菌株生化型存在Ⅲ型(包括生化型Ⅲ-1、Ⅲ-3和Ⅲ-4等亞型)、Ⅱ型和Ⅰ型及无法归属于前人划分的青枯菌生化型标准的非标准生化型,说明百色烟草青枯病菌的生化型结构更加复杂。本研究所有烟区的青枯病菌菌株生化型均包含Ⅲ型和Ⅰ型,唯一的1株Ⅱ型菌株来源于乐业县。值得注意的是,出现了8株非标准生化型菌株,虽然它们出现的区域较窄,只分布在德保县烟区,但有必要监控其在田间的动态消长情况。
我国烟草青枯病抗性种质资源多来自美国和津巴布韦,多为针对演化型Ⅱ型或Ⅲ型菌株选育获得(徐进等,2010)。据王五权等(2015)研究报道,百色烟区的烟草青枯菌在演化型分类上均属于演化型Ⅰ,但目前生产上尚缺乏针对演化型Ⅰ菌株的抗病品种,多年来该烟区尝试利用品种抗病性来控制烟草青枯病为害但收效甚微。因此,今后对百色烟区的抗青枯病种质筛选、评价及布局应针对性地使用当地演化型Ⅰ中的多个强致病力菌株作为测试菌。
本研究的BOX-PCR分析结果与前人关于青枯病菌的遗传多样性与菌株来源具有一定相关性的观点具有一致性(郑向华等,2007;李小杰等,2016),少数电泳图谱基本一致的菌株主要来源于同一烟区。但百色烟草青枯病菌的遗传多样性似乎更复杂,几乎每个菌株都有各自的多样性条带,基于BOX-PCR的聚类分析结果表明所有供试菌株在遗传距离0.60的基础上聚为一大类群;菌株的遗传多态性分析结果与菌株的地理来源、致病力、生化型具有一定的相关性,如少数同源性极高的菌株的地理来源、致病力和生化型均完全一致,但大部分地理来源相同的菌株多态性图谱又存在明显差异,并无明显的地理种群、生化型种群或致病力一致的种群聚在一起。因此,本研究结果仅反映了百色烟区烟草青枯病菌遗传多样性的复杂性,菌株多态性指标并不能显示菌株在致病力、生化型分化上的差异。
4 结论
广西百色烟区烟草青枯病菌具有丰富的遗传多样性和复杂的生化型,使得该病原菌在生存竞争中具有很强的适应能力,给病害的防治带来一定困难,也可能是导致目前该烟区青枯病为害逐年加重的重要原因之一。
参考文献:
蔡刘体,陆宁,沈子霞,石俊雄. 2016. 烟草青枯菌FQY_4宿主特异性候选基因分析[J]. 贵州农业科学,44(10):5-8. [Cai L T,Lu N,Shen Z X,Shi J X. 2016. Analysis of candidate genes of Ralstonia solanacearum FQY_4 for host specificity[J]. Guizhou Agricultural Sciences,44(10):5-8.]
曹毅, 陆宁, 陈兴江, 孟建玉, 商胜华. 2017. 烟草青枯病病圃土壤细菌组成的高通量测序分析[J]. 河南农业科学,46(3):81-85. [Cao Y,Lu N,Chen X J,Meng J Y,Shang S H. 2017. Pyrosequencing analysis of soil bacteria composition in tobacco bacterial wilt disease nursery[J]. Journal of Henan Agricultural Sciences,46(3):81-85.]
岑贞陆, 欧阳秋飞, 谢玲, 陈保善, 朱方容, 林强, 覃丽萍, 蒙姣荣, 史国英, 张雯龙. 2013. 广西桑树细菌性枯萎病病原菌的分离与鉴定[J]. 西南农业学报, 26(3):1054-1057. [Cen Z L, Ouyang Q F, Xie L, Chen B S, Zhu F R, Lin Q, Qin L P, Meng J R, Shi G Y, Zhang W L. 2013. Isolation and identification of pathogenic bacteria from mulberry bacterial wilt in Guangxi[J]. Southwest China Journal of Agricultural Sciences, 26(3):1054-1057.]
亢雅娟, 李双双, 赵丽青. 2008. 植物不同青枯菌生化型的鉴定[J]. 山西师范大学学报(自然科学版), 22(2): 71-75. [Kang Y J, Li S S, Zhao L Q. 2008. Identification on biochemical variants of different Ralstonia solanacearum on plant[J]. Journal of Shanxi Normal University(Natural Science Edition), 22(2): 71-75.]
李小杰, 李淑君, 陈玉国, 王海涛, 李成军, 赵凤霞, 李彦平. 2016. 豫南烟区烟草青枯病菌的致病力及遗传多样性分析[J]. 中国烟草科学, 37(3): 62-66. [Li X J, Li S J, Chen Y G, Wang H T, Li C J, Zhao F X, Li Y P. 2016. Analysis of pathogenicity and genetic diversity of Ralstonia solanacearum on tobacco in southern Henan Province[J]. Chinese Tobacco Science, 37(3): 62-66.]
林海云, 车建美, 刘波, 郑雪芳, 肖荣凤. 2011. 基于BOX-PCR和REP-PCR技术青枯雷尔氏菌遗传多样性分析[J]. 农业生物技术学报, 19(6): 1099-1109. [Lin H Y, Che J M, Liu B, Zheng X F, Xiao R F. 2011. Genetic diversity analysis of Ralstonia solanacearum based on BOX-PCR and REP-PCR[J]. Journal of Agricultural Biotechno-
logy, 19(6):1099-1109.]
王五權, 岑贞陆, 韦忠, 曾泉, 史国英, 周文亮, 农泽梅, 赖洪敏, 黄雪兰, 陆燕飞, 胡春锦. 2015. 广西百色烟草青枯病的危害调查与病原菌分离鉴定[J]. 西南农业学报, 28(6): 2552-2557. [Wang W Q,Cen Z L,Wei Z,Zeng Q,Shi G Y,Zhou W L,Nong Z M,Lai H M,Huang X L, Lu Y F,Hu C J. 2015. Tobacco bacterial wilt survey and pathogenisolation and identification in baise of Guangxi[J]. Southwest China Journal of Agricultural Sciences,28(6): 2552-2557.]
肖田,姚廷山,于庆涛. 2015. 青枯无致病力菌株对烟草青枯病的诱导抗性与控病作用[J]. 西南农业学报, 28(1):207-211. [Xiao T,Yao T S,Yu Q T. 2015. Avirulent strains of Ralstonia solanacearum induced tobacco to resist against bacterial wilt[J]. Southwest China Journal of Agricultural Sciences, 28(1):207-211.]
谢锐鸿, 巫升鑫, 罗正友, 马柱文, 李集勤, 张振臣, 罗静, 张维祥,陈远平, 袁清华. 2014. 粤、闽、贵烟草青枯病菌分离株及其致病力[J]. 微生物学通报, 41(9): 1800-1806. [Xie R H, Wu S X, Luo Z Y, Ma Z W, Li J Q, Zhang Z C, Luo J, Zhang W X, Chen Y P, Yuan Q H. 2014. Pathogenicity of Ralstonia solanacearum isolated from tobacco in Guangdong, Fujian and Guizhou Province[J]. Microbiology China, 41(9): 1800-1806.]
徐进, 顾刚, 潘哲超, 吴畏, 许景升, 张昊, 陈顺辉, 冯洁.2010. 福建烟草青枯菌演化型及生化变种鉴定研究[J].中国烟草学报, 16(6): 66-71. [Xu J, Gu G, Pan Z C, Wu W, Xu J S, Zhang H, Chen S H, Feng J. 2010. Phylotype identification and biovar determination of Ralstonia solanacearum strains isolated from Fujian Province[J]. Chinese Tobacco Science, 16(6): 66-71.]
郑向华, 邓海滨, 刘琼光, 陈泽鹏, 陈永明, 李华平. 2007. 广东省烟草青枯菌的菌系和遗传多样性[J]. 华中农业大学学报, 26(4):463-468. [Zheng X H, Deng H B, Liu Q G, Chen Z P, Chen Y M, Li H P. 2007. Strains and genetic diversity of Ralstonia solanacearum isolated from tobacco in Guangdong Province[J]. Journal of Huazhong Agricultural University, 26(4):463-468.]
Khakvar R. 2009. Diversity and characterization of Ralstonia solanacearum strains in Peninsular malaysia[D]. Serdang: Universiti Putra Malaysia.
Khakvar R, Sijian K, Wong M Y, Radu S, Jones J, Thong K L. 2008. Genomic diversity of Ralstonia solanacearum strains isolated from banana farms in West Malaysia[J]. Plant Patho-
logy Journal, 7(2): 162-167.
Li Y Y, Ji F, Liu H L, Huang J B. 2016. Genetic diversity and pathogenicity of Ralstonia solanacearum causing tobacco bacterial wilt in China[J]. Plant Disease,100(7):1288-1296.
Remenant B, Coupat G B, Guidot A, Cellier G, Wicker E, Allen C, Fegan M, Pruvost O, Elbaz M, Calteau A, Salvignol G, Mornico D, Mangenot S, Barbe V, Médigue C, Prior P. 2010. Genomes of three tomato pathogenswithin the Ralstonia solanacearum species complex reveal significant evolutionary divergence[J]. BMC Genomics,11: 379-394.
(責任编辑 麻小燕)
猜你喜欢
长江蔬菜·学术版(2016年12期)2017-01-12
浙江中西医结合杂志(2017年2期)2017-01-12
现代园艺(2016年3期)2016-03-24
当代化工研究(2016年9期)2016-03-20
天津农业科学(2015年5期)2015-05-30
声屏世界(2014年6期)2014-02-28
西南学林(2013年2期)2013-11-12
中国烟草学报(2012年5期)2012-04-12
中国烟草学报(2012年2期)2012-04-09
