
甘蓝型油菜脯氨酸降解途径关键基因的进化分析
2018-10-11 06:31王翠平陈建伟
江苏农业科学 2018年17期
王翠平, 陈建伟
(宁夏林业研究院种苗生物工程国家重点实验室,宁夏银川 750004
在环境胁迫如干旱、盐、重金属、紫外线等条件下,高等植物可以在短时间内通过迅速大量地积累脯氨酸等渗透调节物质对自身起到保护作用,以此来增强植物对渗透胁迫的抵抗能力[1-5]。在胁迫条件下,脯氨酸可以作为兼容性渗透调节物质、蛋白质和亚细胞结构、活性氧清除剂和氧化还原平衡剂[6-7]。近年来,有研究发现脯氨酸参与了开花及胚胎发育过程[8-11]。高等植物中,吡咯啉-5-羧酸合成酶(P5CS)是脯氨酸合成途径的关键酶[8-9],而脯氨酸脱氢酶(proline dehydrogenase,PDH)是脯氨酸降解途径的关键酶[10]。在拟南芥中,PDH定位于线粒体,由2个基因编码(AtPDH1,At3g30775;AtPDH2,At5g30870),脯氨酸处理会诱导PDH1表达,而胁迫处理抑制PDH1表达[11]。
植物进化过程中多倍化是重要的进化动力,并且许多作物经历了多倍化过程,多倍体能够为新表型的出现提供原材料,具有进化上的优势[12]。多倍化之后由于染色体重组和基因重排,有一些基因表达模式会发生变化,甚至会发生丢失[13-14]。芸薹属(Brassica)植物是研究由于多倍化引起的基因缺失、沉默和偏向表达的非常好的模式植物体系,禹氏三角模型能够很好地解释芸薹属植物的进化关系[15]。甘蓝型油菜(Bassicanapus)是最新形成的异源四倍体,来源于2个二倍体白菜(B.rapa)和甘蓝(B.oleracea)种间杂交,并且经历了染色体加倍的过程[16]。而白菜、甘蓝和拟南芥起源于共同的祖先,大概在1 450万~2 040万年之前这个祖先分化为拟南芥和白菜、甘蓝的祖先[17]。比较物理图谱结果表明,白菜和甘蓝的祖先经历了三倍化和基因重排的过程[18]。且白菜基因组测序数据也表明,白菜中大多数基因具有2~3个同源基因[19]。那么,推测异源四倍体油菜中同源基因拷贝数应该更多。有报道称,油菜中基因拷贝数目可能由于进化过程中基因丢失的原因而减少[20]。
胁迫条件下脯氨酸积累是一个非常保守且有效的过程,然而迄今为止仍不清楚四倍体中脯氨酸代谢相关基因的进化模式及其与二倍体祖先脯氨酸代谢相关基因的关系。之前研究发现,油菜中含有6个脯氨酸合成途径关键基因P5CS1,即该基因没有发生丢失,并且在不同器官中与胁迫诱导情况下不同来源的P5CS1基因表达模式不同[21]。本研究集中探讨了甘蓝型油菜中脯氨酸降解代谢相关基因PDH1和PDH2的进化命运,包括二倍体亲本进化上的来源、与二倍体亲本物种的基因序列差异和重复基因是否发生丢失。研究结果能够加深多倍化对脯氨酸代谢相关基因影响的理解,为多倍体表型和重复基因调控的关系提供理论依据。
1 基因序列分析
白菜和甘蓝的相应基因序列由中国农业科学院蔬菜花卉研究所王晓武研究员提供,并且通过数据库检索和比对(http://www.brassica.info/; http://www.ocri-genomics.org/bolbase/index.html)。油菜基因序列来自于开放的油菜基因组序列网站(http://www.genoscope.cns.fr/brassicanapus/)。核酸序列分析应用DNAMAN软件(Lynnon Biosoft,Vaudreuil,Quebec,Canada)。通过对同源基因对应位置的部分基因组DNA进行比对和序列分析,应用MEGA4软件中邻位相连法构建进化树[22]。
2 结果与分析
2.1 油菜中脯氨酸合成代谢相关重复基因
2.1.1 白菜、甘蓝、油菜中的PDH1基因概况 序列分析结果表明,白菜中存在3个PDH1的同源基因(http://www.brassica.info/)(ChrA2,Bra020731;ChrA6,Bra025421;ChrA9,Bra036196),甘蓝中存在3个PDH1基因(http://www.ocri-genomics.org/bolbase/index.html)(Bol033129;Bol043056;Bol032297),甘蓝型油菜中存在5个PDH1基因(http://www.genoscope.cns.fr/brassicanapus/) (ChrC7,NC_027773.1;ChrC5,NC_027771.1;ChrC5,NC_027771.1;ChrA9,NC_027765.1;unplaced genomic scaffold,NW_013650408.1)。根据系统命名法则[23],将白菜的3个PDH1同源基因分别命名为BraA.PDH1.a、BraA.PDH1.b、BraA.PDH1.c;将甘蓝的3个PDH1基因分别命名为BolC.PDH1.a、BolC.PDH1.b、BolC.PDH1.c;将甘蓝型油菜的5个PDH1基因分别命名为BnaA.PDH1.a、BnaA.PDH1.b、BnaC.PDH1.c、BnaC.PDH1.d、BnaC.PDH1.e。与拟南芥相同,芸薹属植物的PDH1基因也具有4个外显子和3个内含子,除第1外显子比拟南芥第1外显子少3个碱基以外,其他外显子碱基数目一致(表1)。芸薹属植物白菜、甘蓝和甘蓝型油菜的基因组序列(从ATG到终止密码子)长度为2 201~2 671 bp,然而编码区(CDS)长度一致,均为1 497 bp,所编码的蛋白质都含有498个氨基酸。

表1 芸薹属PDH1基因结构对比
2.1.2 白菜、甘蓝和甘蓝型油菜中PDH2基因概况 基因组测序结果表明,白菜中存在1个PDH2基因(Bra028202),位于白菜A4号染色体上,命名为BraA.PDH2.a;甘蓝中存在1个PDH2基因,命名为BolC.PDH2.a;甘蓝型油菜中存在2个PDH2基因,分别命名为BnaA.PDH2.a、BnaC.PDH2.b。芸薹属植物PDH2基因cDNA编码区长度为1 431 bp,含有4个外显子3个内含子,和拟南芥一致(表2)。

表2 芸薹属PDH2基因结构对比
2.2 脯氨酸合成酶基因的同源性和进化关系
2.2.1PDH1基因的同源性和进化关系 为研究甘蓝型油菜中的5个PDH1基因的二倍体祖先在甘蓝型油菜染色体加倍过程中来自于白菜基因组还是甘蓝基因组,将5个基因分别与白菜及甘蓝的PDH1基因的基因组序列及编码区序列做两两序列比对,结果如表2所示。同时,将甘蓝型油菜、白菜、甘蓝和拟南芥的同源PDH1基因的基因组DNA序列作多重比对,构建该基因的系统发生树(图1)。
序列相似性分析结果(表3)表明,BnaA.PDH1.a和BraA.PDH1.a、BnaA.PDH1.b和BraA.PDH1.c、BnaC.PDH1.c和BolC.PDH1.a、BnaC.PDH1.d和BolC.PDH1.b、BnaC.PDH1.e和BolC.PDH1.c之间的基因组DNA相似性分别为96.34%、99.67%、99.78%、98.48%、91.62%,而它们之间编码区相似性更高,分别为99.20%、99.67%、99.80%、99.20%、99.53%,无论是基因组序列还是编码区序列均高于同系列其他比对。因此,推断BnaA.PDH1.a、BnaA.PDH1.b、BnaC.PDH1.c、BnaC.PDH1.d和BnaC.PDH1.e可能分别来

源于BraA.PDH1.a、BraA.PDH1.c、BolC.PDH1.a、BolC.PDH1.b和BolC.PDH1.c。而白菜的BraA.PDH1.b在甘蓝型油菜中发生丢失。
而基于相对应的基因组序列的进化关系分析表明,BnaA.PDH1.a、BnaA.PDH1.b、BnaC.PDH1.c、BnaC.PDH1.d、BnaC.PDH1.e分别为白菜BraA.PDH1.a、BraA.PDH1.c和甘蓝BolC.PDH1.a、BolC.PDH1.b、BolC.PDH1.c的直向同源基因(图1)。
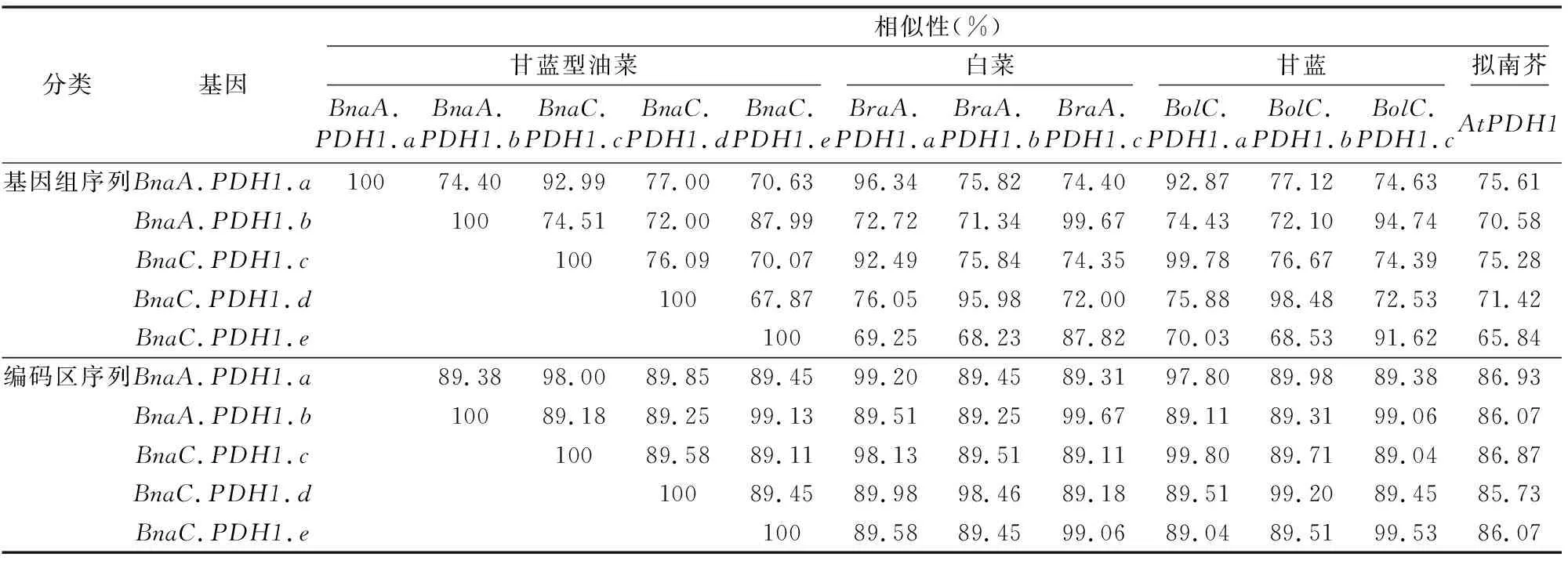
表3 甘蓝型油菜PDH1基因的基因组序列和编码区序列与二倍体亲本白菜和甘蓝PDH1基因序列相似性分析
2.2.2PDH2基因的同源性和进化关系 甘蓝型油菜基因组中含有2个同源的PDH2基因,为了研究这2个基因的祖先来源,将2个基因分别与白菜及甘蓝的PDH2基因基于基因组序列和编码区序列做两两序列比对,比对结果如表4所示。同时,将甘蓝型油菜、白菜、甘蓝和拟南芥的所有已知序列的同源PDH2基因的基因组DNA序列作多重比对,构建该基因的系统发生树(图2)。
序列相似性分析结果表明,BnaA.PDH2.a与BraA.PDH2.a、BnaC.PDH2.b与BolC.PDH2.a基因组DNA序列相似性分别高达96.73%、99.67%(表4),而它们之间编码区相似性更高,分别为99.86%、99.93%(表4),无论是基因组序列还是编码区序列均高于同系列其他比对,推测BnaA.PDH2.a、BnaC.PDH2.b可能分别来源于BraA.PDH2.a、BolC.PDH2.a。
且基于相对应的基因组序列的进化关系分析也表明,BnaA.PDH2.a、BnaC.PDH2.b分别为白菜BraA.PDH2.a、甘蓝BolC.PDH2.a的直向同源基因(图2)。


表4 甘蓝型油菜中PDH2基因的基因组片段与二倍体祖先中相对应的基因核苷酸序列相似性
3 讨论
3.1 油菜的PDH1基因可能发生了1个拷贝的丢失
甘蓝型油菜(B.napus)起源于白菜(B.rapa)和甘蓝(B.oleracea)的杂交,并且经过了染色体加倍,而其祖先白菜和甘蓝自身也经历了基因组三倍化和重排的过程[16],这样推测甘蓝型油菜中每个基因同源基因的拷贝数最多可以达到6个。然而研究发现,由于基因丢失现象的存在,甘蓝型油菜中同源基因的拷贝数平均为4个[20]。研究表明,甘蓝型油菜中存在3个ALCATRAZ(ALC)基因[24]、3个GPAT4(sn-glycerol-3-phosphate acyltransferase 4)基因,然而存在的6个PSY(Phytoene Synthase)基因中有3个来源于白菜,另外3个来源于甘蓝[25],而笔者之前的研究发现,甘蓝型油菜中存在6个P5CS1基因[21]。本研究中,甘蓝型油菜中存在5个PDH1基因,而其祖先白菜和甘蓝中各存在3个PDH1基因,进化过程中甘蓝型油菜可能发生了PDH1基因1个拷贝的丢失,而根据序列比对和进化分析结果,推测来自于二倍体祖先白菜的PDH1基因的1个拷贝发生丢失。而甘蓝型油菜中存在2个PDH2基因,其祖先白菜和甘蓝中各存在1个PDH2基因,甘蓝型油菜中PDH2基因没有发生丢失。
3.2 油菜中脯氨酸代谢相关基因与二倍体亲本来源基因高度同源
根据白菜和甘蓝全基因组测序结果,白菜和甘蓝中PDH1和PDH2同源基因拷贝数都分别为3、1,而甘蓝型油菜中拷贝数为5、2。序列比对结果表明,甘蓝型油菜脯氨酸代谢途径相关基因均与其亲本白菜和甘蓝来源的基因高度同源,PDH1和PDH2同源基因和亲本来源基因的基因组DNA同源性分别为91.62%~99.78%、96.73%~99.67%。说明这些基因在甘蓝型油菜进化过程中高度保守。这与之前的甘蓝型油菜基因克隆的相关研究报道一致,甘蓝型油菜中克隆得到3个PISTILLATA基因,两两序列相似性为96.49%~98.72%[26]。脯氨酸合成途径上不同基因拷贝数的不同或许说明了这些基因在进化上的重要程度。
4 结论
以甘蓝型油菜、白菜和甘蓝为试验材料,研究了甘蓝型油菜中脯氨酸代谢途径同源基因的进化命运,探讨了多倍化对同源基因进化的影响。主要结论如下:通过白菜、甘蓝和甘蓝型油菜的基因组数据库搜索和序列比对,确定了脯氨酸降解途径关键基因(PDH1、PDH2)在甘蓝型油菜中均存在多个拷贝,序列比对结果表明,它们和二倍体亲本白菜及甘蓝的对应基因高度同源;和亲本相比,甘蓝型油菜的PDH1基因来自于二倍体祖先甘蓝的1个拷贝发生了丢失,而PDH2基因没有发生丢失。
猜你喜欢
华人时刊(2023年1期)2023-03-14
汉字汉语研究(2021年2期)2021-08-30
西藏农业科技(2019年3期)2019-11-04
汉字汉语研究(2019年2期)2019-08-27
西藏农业科技(2019年1期)2019-07-25
西藏农业科技(2018年4期)2018-04-25
现代农业科技(2017年8期)2017-06-10
河北书画研究(2016年3期)2016-04-28
西南农业学报(2016年6期)2016-04-16
中国果业信息(2015年6期)2015-01-23
