
泰万菌素的抗炎效果分析
2019-01-16 12:19李仕超
猪业科学 2018年12期
文 丽,李仕超(译)
(上海伊克拜克兽药销售有限公司,上海 200333)
0 引言
炎症的消除对于机体在感染或组织损伤后平衡的恢复至关重要。例如,支原体和胸膜肺炎放线杆菌定植在猪肺中,诱导产生白细胞毒素,裂解中性粒细胞和巨噬细胞。这可以促进自发的炎症反应,活化的白细胞释放更多的促炎因子,包括IL-1α,IL-1β, IL-6, CXCL-8 (即IL-8)和白细胞三烯B4。这种自我放大反应正是肺炎病理学特征。
嗜中性粒细胞的凋亡对于炎症消除必不可少,以质膜出泡、染色体收缩、DNA破碎及凋亡小体的形成为特征。如果清除不彻底,凋亡小体会造成组织器官继发性坏死。巨噬细胞消除凋亡小体,即胞葬作用,在炎症的消除中扮演关键角色。胞葬作用可以诱导抗炎巨噬细胞表型,降低促炎因子,如IL-6,LBT4,CXCL-8和TNF-α;促进抗炎因子,如TGF-β,LXA4和IL-10。消炎因子包括脂氧素类和消退素类,可以帮助消炎。脂氧素类有抗炎和消炎双重作用如增强胞葬作用,诱导抗炎因子IL-10的产生,减少致炎因子,起到镇痛作用。这个脂质介质家族由花生四烯酸通过15-脂氧合酶和15(S)-HETE前体产生。二十二碳六烯酸代谢物如消退素D1(RvD1)是由omega-3和6脂肪酸合成而来。RvD1减少嗜中性粒细胞在肺部聚积,缓解水肿,抑制局部促炎因子的产生。RvD1和它的阿司匹林诱生型差向异构体,在革兰氏阴性细菌肺炎自我消炎中起关键作用。在过去10年里,兽药和人药领域的研究者们一直在探索新疗法,该种方法通过刺激抗炎或消炎因子来阻止机体的自我延续性炎症反应。
大环内酯类抗生素有广泛的免疫调节属性,它可以在巨噬细胞内聚集,胞内浓度可以比外界高500倍,给予了这些药物卓越的药效。关于大环内酯类怎样改变发炎组织中脂质介质的产生,我们却知之甚少。为此,我们使用了猪细胞模型和泰万菌素——一种新型广谱抗菌药,是泰乐菌素的衍生物。泰万菌素是从泰乐菌素经3-乙酰基-4异戊酰基修饰后而来的第3代大环内酯类广谱抗生素(酒石酸乙酰基异戊酰基泰乐菌素)。泰万菌素现在用于控制鸡和猪由支原体等病原引起的呼吸道疾病和肠道细菌感染。最新研究发现,鼠或猪感染猪繁殖与呼吸障碍综合征病毒(PRRSV)后由脂多糖(LPS)引起的肺部炎症反应模型中,泰万菌素可以通过抑制NF-κB起到抗炎作用,NF-κB是可以介导促炎因子如CXCL-8(即IL-8)合成的转录因子。使用泰万菌素和猪白细胞作为模型,目前研究表明除了抗菌作用之外,这种抗生素还可以促使嗜中性粒细胞凋亡、起到胞葬作用,抑制促炎因子LTB4的产生,促进LXA4和RvD1的合成。
1 试验结果
1.1 泰万菌素诱导猪嗜中性粒细胞浓度和时间依赖性凋亡
Caspase-3裂解,在细胞凋亡中引起DNA断裂,泰万菌素处理猪嗜中性粒细胞中明显高于对照组(图1)。通过原位末端标记染色法和ELISA检测法对细胞凋亡产生的单聚和寡聚核苷酸进行检测,证实了泰万菌素诱导嗜中性粒细胞的凋亡作用有浓度和时间依赖性(0.5 h和1 h)(图1)。林可霉素是另外一种猪肺炎的治疗用抗生素,不会诱导细胞凋亡。该试验也检测了阳性对照staurosporine组对细胞凋亡的诱导作用。
图1A:凋亡的嗜中性粒细胞末端标记法的荧光图。
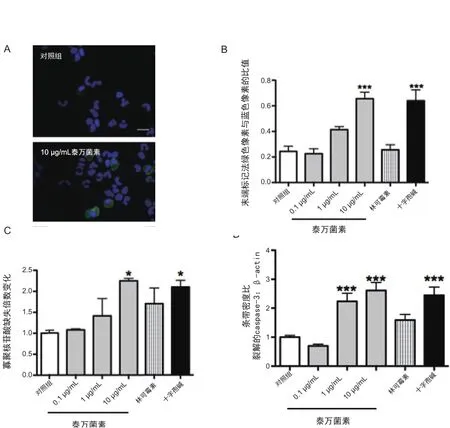
图1 泰万菌素诱导猪嗜中性粒细胞的凋亡
绿色是末端标记阳性,蓝色是DAPI染色的细胞核。泰万菌素处理组(1 h, 10 µg/mL)的嗜中性粒细胞末端标记阳性数量(凋亡)更多,1000×放大。
图1B:1 h处理后末端标记试验的定量。
数值为末端标记法绿色像素与蓝色像素的比值。数值是指平均值±(n=3~6/组) ***P<0.001(与对照组差异显著)。
图1C:ELISA试验结果显示细胞凋亡时释放的单聚和寡聚核苷酸。
数值是指对照组吸光度的倍数变化。数据是各实验组溶液处理1 h后获得:对照组(含酚红的10%胎牛血清),泰万组、阳性组(staurosporine)、林可霉素组使用不同浓度。数值是指平均值±SEM(n=3~4/组) *P<0.05。
图1D:猪嗜中性粒细胞处理0.5 h后,促凋亡的caspase-3相对于β-actin的密度分析。
对照组(含酚红的10%胎牛血清)、泰万组(0.1, 1.0或10 µg/mL)、阳性组(10-6mol/L staurosporine)、林可霉素组(11.3×10-6mol/L)。数值是裂解的caspase-3((17 kDa)相对于β-actin(45 kDa)的条带密度,表示为与对照组的比率。数值是组平均值±SEM(n=3~6/组). ***P<0.001(与对照组差异显著)。
1.2 泰万菌素诱导猪单核细胞源的巨噬细胞浓度和时间依赖性凋亡
泰万菌素处理12 h和14 h后也可以诱导猪单核细胞源的巨噬细胞浓度和时间依赖性凋亡, 更早的时间点则不能(图2)。林可霉素也不能。这个用原位末端标记法、单聚和寡聚核苷酸的产生、裂解caspase-3的超标也得到证实。这些影响在阳性对照组(staurosporine)中也可以看到,林可霉素组不能。
单核细胞源的巨噬细胞用以下溶液处理:对照组(10%胎牛血清培养基)、泰万组(0.1, 1, 或 10µg/mL)、阳性组(10-6mol/L staurosporine)、林可霉素组(11×10-6mol/L)。
图2A: 泰万菌素对巨噬细胞凋亡的诱导作用呈时间和浓度依赖性。

图2 泰万菌素时间和浓度依赖性诱导猪巨噬细胞的凋亡
数值是每张图中末端标记的细胞与核DAPI染色(0.5-24 h)的比值。数值是指平均值(n=4-8/组) *P<0.05, ***P<0.001。
图2B/C: 同样条件下,凋亡细胞ELISA试验结果如图2B(12 h)和图2C(24 h)所示。
试验是诱导细胞凋亡后对细胞质中单聚和寡聚核苷酸的定量检测。数值是指对照组吸光度的倍数。数值是指平均值±SEM (n=3~5/组). *P<0.05, **P<0.01。
D/E图: 泰万菌素活化猪巨噬细胞中的caspase-3如图D(12 h)和图E(24 h)所示。
数值是指平均值±SEM (n=3~5/组). *P<0.05.
1.3 泰万菌素不影响猪嗜中性粒细胞和巨噬细胞的坏死
泰万菌素诱导嗜中性粒细胞凋亡的时间点(0.5 h和1 h,图1)或诱导巨噬细胞凋亡的时间点(12 h和24 h,图2),通过乳酸脱氢酶(LDH)释放的检测发现,抗生素不会改变这些细胞的坏死(图3)。

图3 泰万菌素在体外对静息细胞坏死无影响
通过对细胞培养上清液中LDH蛋白的定量来检测分泌型乳酸脱氢酶(LDH),从而评估细胞坏死。猪嗜中性粒细胞用以下溶液孵育:对照组(含酚红的10%胎牛血清)、泰万组(10µg/mL)、林可霉素组(11×10-6mol/L,等摩尔数)、阳性对照组(1% Triton X-100)图3 A(0.5 h)和图3B(1 h):数值是指对照组的倍数,数值是指平均值 ± SEM (n = 9), **P<0.01。图3C :猪巨噬细胞的细胞坏死(通过LDH的释放来检测)。
分别用以下溶液孵育0.5-24 h:对照组(10%胎牛血清培养基)、泰万组(0.1, 1.0或 10 µg/mL)、阳性对照组(1% Triton X-100)。数值是指对照组的倍数,对照组与泰万组之间数值差异不大(n=6~9/组),数值是指平均值 ± SEM,*P< 0.05。
1.4 泰万菌素通过巨噬细胞促进嗜中性粒细胞的胞葬作用,不会改变吞噬作用
为了检测泰万菌素诱导嗜中性粒细胞的凋亡是否导致胞葬作用,猪嗜中性粒细胞用泰万菌素(0.1, 1.0或10 µg/mL)或上述对照组溶液孵育0.5 h,再与单核细胞源的巨噬细胞共同孵育2 h。泰万菌素浓度依赖性显著增加胞葬作用(图4A)。诱导巨噬细胞的胞葬作用与巨噬细胞吞噬作用之间没有关联(通过酵母聚糖颗粒的吸收检测),表明在凋亡小体中,泰万菌素诱导的巨噬细胞摄入底物的激活作用至少是在一定程度上是有选择性的(图4B)。
图4A:过氧化物酶(MPO)是中性粒细胞的标记物。
在嗜中性粒细胞用泰万菌素(0.1, 1, 10 µg/mL)处理0.5 h后的巨噬细胞中发现MPO的浓度增加。未处理的嗜中性粒细胞处理2 h(10% FBS 0.5 h)处理的猪巨噬细胞作为对照。
数值用对照组MPO活性测量值的倍数表示,数值是平均值±SEM (n=3/组). ***P<0.001.
图4B:酵母聚糖颗粒的甘露糖依赖性吞噬作用在猪巨噬细胞中没有变化,2 h时胞葬作用增强。
单核细胞源的巨噬细胞用1 mg/mL的酵母聚糖颗粒孵育2 h (n=6/组),对照组10% FBS,泰万组10µg/mL泰万菌素孵育2 h。含有一个或多个酵母聚糖的巨噬细胞计数为阳性细胞,数值是阳性巨噬细胞与总数的比值,数值为平均值±SEM。

图4 泰万菌素促进胞葬作用,但在猪体外单核细胞源巨噬细胞中不调节甘露糖依赖性的吞噬作用
1.5 泰万菌素抑制嗜中性粒细胞中促炎因子LTB4的产生
通过超高温液相色谱质谱法(UHPLC-MS)检测发现,泰万菌素抑制钙离子通道活化的嗜中性粒细胞中LTB4的产生(图5)。钙离子通道的活化反应使嗜中性粒细胞高度活化,随后升高的LTB4,是在发炎组织中合成。
图5A:通过反相-高效液相色谱法(RP-HPLC)检测猪始嗜中性粒细胞中的促炎因子LTB4分泌水平。
对照组用 10% HI-FBS in HBSS处理,试验组10 µg/mL泰万菌素、加或不加3µM 钙离子载体 A23187 ,0.5 h。数值代表LTB4峰面积的倍数,数值是平均值±SEM (n=7~10/组).**P<0.01(与对照组); #P< 0.05(3 µM A23187)。
图5B:猪嗜中性粒细胞用以下处理0.5 h后,检测LTB4水平。通过液相色谱质谱法(LC-MS),数值代表LTB4浓度,数值是平均值±SEM (n=10~15/ 组). ***P<0.001(与对照组); ##P< 0.01(3 µM A23187)。
1.6 泰万菌素LPS刺激的巨噬细胞中抑制促炎因子CXCL-8和IL-1α的释放
相对于对照组,泰万菌素处理组中,LPS刺激的巨噬细胞产生的CXCL-8(图6A)和IL-1α(图6B)明显更低。
(图6B)明显更低。在有或无LPS(1 µg/mL)处理2 h前提下,珠状荧光复合系统被用于评估对照组或泰万菌素(10 µg/mL)处理组的猪巨噬细胞CXCL-8和IL-1α水平。
猪巨噬细胞用10% HI-FBS或10 µg/mL泰万菌素,在有或无LPS(1 µg/mL)条件下处理2 h后收集上清液,用于分泌型趋化因子/细胞因子的检测。使用珠状荧光试验和标准曲线,检测了分泌蛋白量,数值单位 pg/mL。
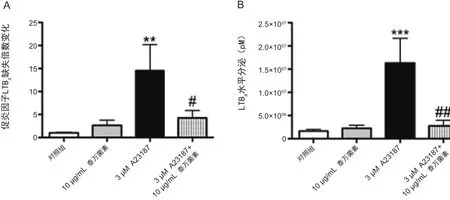
图5 泰万菌素抑制钙离子通道活化的嗜中性粒细胞中促炎因子LTB4的产生

图6 泰万菌素处理抑制CXCL-8和IL-1α在LPS刺激的巨噬细胞中的分泌
在LPS刺激的巨噬细胞中,CXCL-8(图6A)和IL-1α(图6B)的蛋白分泌受到泰万菌素抑制。数值代表平均值±SEM (n=3~4/组). *P<0.05, **P< 0.01(未刺激组); #P<0.05 (LPS刺激的巨噬细胞)。
1.7 泰万菌素促进嗜中性粒细胞中消炎因子LXA4和RvD1的释放
使用液相色谱/质谱法(LC/MC)评估猪嗜中性粒细胞(经对照组溶液或泰万菌素孵育)LXA4和其前体15S- HETE,以及RvD1的产生。泰万菌素促进LXA4(图7A)和RvD1(图7C)的释放。15S- HETE的产生值,未达到具有统计学意义的水平(图7B)。
用10% HI-FBS或10µg/mL泰万菌素处理0.5 h,用LC/MC检测:LXA4的分泌水平(图7A);
前体15S-HETE的分泌水平(图7B);RvD1的分泌水平(图7C)。数值分别通过与相应物质洗脱峰面积标准曲线计算得来。数值代表平均值±SEM (n=18~20). *P<0.05, ***P<0.01 (未刺激对照)。
1.8 泰万菌素在嗜中性粒细胞中上调胞浆型磷脂酶A2(cPLA2)和磷脂酶C(PLC)的活性
为了更进一步探究泰万菌素对嗜中性粒细胞脂质调节介质影响的机制,又进行试验对胞浆型磷脂酶A2和磷脂酶C(PLC)的活性进行了评估。cPLA2可以从膜磷脂释放花生四烯酸,花生四烯酸可以由甘油二酯脂肪酶代谢产生,三磷酸肌酐可以由膜磷脂PLC产生。这种花生四烯酸的形成可以从释放的花生四烯酸中合成脂质介质。与对照组相比,在泰万菌素孵育的猪嗜中性粒细胞中,胞浆型磷脂酶A2(cPLA2)和磷脂酶C(PLC)的活性显著提高(图8)。
图8A:胞浆型磷脂酶A2(cPLA2)活性在泰万处理组(10 µg/mL,0.5 h)和对照组(10% HI-FBS)无差异。数值代表 cPLA2活性(µmol/min/mL),数值代表平均值± SEM (n=5/组). *P>0.05.

图7 泰万菌素促进消炎因子LXA4和RvD1的分泌

图8 泰万菌素在猪嗜中性粒细胞中促进细胞内磷脂酶PLC活性
图8B:泰万处理组(10 µg/mL ,0.5 h)胞内磷脂酶PLC活性显著高于对照(HBSS中10% HI-FBS)。数值代表PLC酶活性,是对照组的倍数。数值代表平均值±SEM (n=4/组). *P<0.05.
2 讨论
细菌感染治疗中,若提供抗菌和抗炎/消炎双重效果,可产生优良的临床效益。该研究使用泰万菌素在猪嗜中性粒细胞和巨噬细胞中诱导免疫调节这一模型,展示了一种新型的抗生素诱导嗜中性粒细胞释放有效的消炎因子RvD1和 LXA4,这种作用与PLC活性有关。研究发现泰万菌素可以抑制促炎因子的产生,如嗜中性粒细胞中的LTB4和LPS刺激的巨噬细胞中的CXCL-8和IL-1α。最终泰万菌素在嗜中性粒细胞和单核细胞源的巨噬细胞中诱导浓度和时间依赖性凋亡,促进胞葬作用。至少在林可霉素(另一种猪肺炎的轮换治疗药物)没有这种作用。总之,这项研究表明泰万菌素,除了其抗菌活性以外,还有显著的免疫调节特性,为体内治疗炎症提供了极大可能性,有待进一步研究。猪细菌性肺炎中的病理变化和急剧的器官损伤,至少可以减少局部细胞坏死和嗜中性粒细胞的自我放大作用,这和动物、人类多种炎性疾病类似。这些垂死细胞的控制,通过凋亡和胞葬作用的清除,代表了炎症消除的关键阶段。在机体平衡条件下,嗜中性粒细胞是没有活性的,通过凋亡(细胞程序性死亡)在炎症位点被移除。在细胞凋亡过程中,在细胞质膜完整性存在的情况下,细胞器被摧毁,从而阻止促炎因子和组织溶解复合物释放到周围环境中。由此而论,白细胞凋亡的促进作用被认为赋予了药物显著的消炎属性,这种生理调控程序不影响其抗菌功能。近期有研究发现,细胞周期蛋白依赖的激酶抑制剂与大环内酯类抗生素,如替米考星和托拉霉素一样,通过诱导嗜中性粒细胞凋亡来增强消炎作用。与之一致,该研究也是基于抗生素促进炎症细胞凋亡的设想,其可能对于宿主炎症为主要发病机制的传染性疾病中有重要治疗效果。近期研究表明:泰万菌素在鼠科急剧肺损伤模型中,以及在用猪蓝耳病毒攻毒的猪只中通过抑制NF-κB途径的激活,起到免疫调节作用。这些研究可能指向与白细胞内高亲嗜性利用这种复合物相关的胞内机制。类似于这种设想,托拉霉素及其他大环内酯类抗生素的胞内聚积可能抑制胞内磷脂酶(调控脂质介质合成和磷脂依赖性细胞凋亡),泰万菌素确实可以降低磷脂酶A2的表达。与之对应,这项研究表明,在猪嗜中性粒细胞中,泰万菌素可能增加细胞质内PLC和磷脂酶A2。这种矛盾是否反应了不同模型中不同的生物功能,还有待进一步研究。
除了抗菌效果之外,大环内酯类抗生素可能调节宿主白细胞反应,引起血管生成作用和抗癌作用。与这些结果类似,该研究报道了泰万菌素抑制巨噬细胞中的促炎因子IL-1α和趋化因子CXCL-8。关于这些药物是如何影响脂质介质的产生,目前还知之甚少。LTB4和CXCL-8一样是与肺炎发病有关的强大的中性粒细胞趋化因子。LTB4还与嗜中性粒细胞脱粒、超氧化物自由基和弹性蛋白酶的局部释放有关,与β-整合素一起,引起黏膜群集表型,并进一步维持白细胞招募和炎性损伤。该研究证实泰万菌素除了抗菌活性以外,还可以直接抑制LTB4从激活的猪嗜中性粒细胞中释放。
该研究通过超高效液相色谱-质谱法(UHPLCMS),鉴定了一种新型抗生素的效果——触发产生有效的消炎脂质代谢物RvD1和LXA4。近期报道了托拉霉素可以通过猪嗜中性粒细胞促进LXA4的产生。通过不断进行的有关LXA4用于人类齿龈炎治疗、Ⅱ型糖尿病、动脉粥样硬化和哮喘临床试验,这些消炎脂质介质的临床意义正在不断得到人们重视。此外,鉴于LTB4的同时抑制作用,泰万菌素可以为研究抗生素诱导的脂质介质类型转换提供有力的武器,对于炎症的消除至关重要。胞葬作用确实促进脂质介质类型转换。在这个过程中,嗜中性粒细胞在前列腺素E2和D2的作用下,基因表达水平上发生变化,通过增加15-脂氧合酶的表达,产生脂氧素和D类消退素,从而开启花生四烯酸的代谢。泰万菌素是否可以改变脂质介质类型转换相关酶的转录,或者泰万菌素是否作用于变构位点来实现这些酶的抑制或刺激,还有待进一步研究。该研究尚且是体外试验,还需要猪只活体感染模型来证实泰万菌素的这种作用,有可能为大环内酯类抗生素乃至其他药物的新促分辨特征铺平道路。这些在抗生素耐药性的环境中会发挥什么作用,也是未来研究的一个重要方向。
猜你喜欢
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
昆明医科大学学报(2021年12期)2021-12-30
农业与技术(2021年21期)2021-11-17
皮肤病与性病(2021年3期)2021-07-30
昆明医科大学学报(2021年2期)2021-03-29
家庭影院技术(2018年9期)2018-11-02
中国制笔(2017年2期)2017-07-18
米娜·女性大世界(2016年8期)2016-08-17
中国感染与化疗杂志(2015年5期)2015-01-23
食品工业科技(2014年13期)2014-03-11
