
不同抗性烟草品种內源激素对赤星病胁迫的响应差异
2021-01-18 19:40杨志晓王轶谢升东郑少清夏海乾陈懿余世洲王自力曹毅王仁刚董祥立李治模张维军林英超
中国烟草科学 2021年2期
杨志晓 王轶 谢升东 郑少清 夏海乾 陈懿 余世洲 王自力 曹毅 王仁刚 董祥立 李治模 张维军 林英超
摘要:为进一步明确烟株内源激素与赤星病抗性的关系,以抗赤星病烟草品种净叶黄和感病品种长脖黄为研究对象,通过盆栽种植并接种赤星病菌,比较不同抗性品种内源激素含量对赤星病胁迫的响应差异。研究结果显示,在赤星病胁迫下,净叶黄、长脖黄的脱落酸(ABA)含量和乙烯(ET)释放量均呈上升趋势,而生长素(IAA)以及细胞分裂素(CIK)含量表现为降低。同时,赤星病胁迫导致净叶黄的赤霉素(GA)、茉莉酸(JA)含量增加,引起长脖黄的JA含量降低,GA含量则先增加后降低。与净叶黄相比,长脖黄的ABA含量增幅较小,而GA、IAA、CTK与JA含量降幅较大,并且ET释放量较高。综上,在赤星病胁迫下,内源激素的含量差异以及变化幅度可以反映不同品种的抗病能力。
关键词:赤星病胁迫;内源激素;烟草;品种;抗病性
植物体中存在的内源激素主要包括脱落酸、乙烯、生长素、细胞分裂素以及赤霉素等不同类别物质,每一种类的激素都在一定浓度范围对植株的生长发育进程产生调控作用。植物内源激素可以参与抵御病原菌侵染,诱导寄主的防御反应,避免或者械轻病原菌所造成的危害,与抗病性存在紧密联系。徐以华等指出,在水稻与穗枯病菌互作反应中,穗枯病菌能够诱导一系列内源激素含量发生改变,且这种诱导效应在抗病品种中更为明显。钟丽娟等研究发现,烟株在感染马铃薯Y病毒后,IAA含量显著降低,且感病品种的下降幅度大。在灰斑病菌作用下,大豆叶片的IAA含量升高有利于品种抗病能力增强,但感病品种的IAA、GA含量却出现降低。基于此,深入分析内源激素在植物抵御病原菌侵染过程中的作用对于病害防治工作具有重要的指导价值。
烟草赤星病是由链格孢菌Alternaria alternate(Fries)Keis引起的一种真菌性病害,能够对烟叶生产造成巨大损失。目前有关烟草受到赤星病侵染后其内源激素的变化规律尚未见报道。因此,本研究选择2个具有不同赤星病抗性的烟草品种作为试验材料,分析抗、感病品种在赤星病胁迫条件下其内源激素包括脱落酸(ABA)、茉莉酸(JA)、生长素(IAA)、赤霉素(GA)、细胞分裂素(CK)含量与乙烯(ET)释放量的差异,旨在探讨烟草内源激素与赤星病抗性的关系,为抗病烟草品种培育与病害的科学防治提供理论参考。
1材料与方法
1.1试验材料
供试材料分别为高抗、高感烟草赤星病烟草品种净叶黄(JYH)、长脖黄(CBH)。JYH是由CBH的抗赤星病单株经系统选育而成,该品种为我国抗赤星病育种的主要抗源,利用其作为亲本已培育出多个高抗或耐病品种。
选择具较高致病性的烟草赤星病菌种GM831
做为人工接种使用的赤星病菌株,该菌株由贵州省烟草科学研究院植保团队提供。
内源激素ABA、GA、IAA、CTK和JA所用检测标准样品为Sigma公司产品。
1.2试验方法
2020年在贵州省烟草科学研究院开展盆栽试验。漂浮育苗,在6叶1心时选择生长整齐、均匀一致的壮苗移栽至装有10kg土的聚乙烯盆中,每盆内植烟1株,盆栽的行距、株距按照1.1m、0.55m排列。每个品种重复3次,每重复种植50株。盆栽壤为壤质潮土(有机质8.52gkg、全氮0.89gkg、速效氮66.46mgkg、速效磷24.43mgkg、速效钾108.15mgkg、pH792)。每盆烟株所用N、P、K的施入量依次是0.13、0.13以及0.39gkg,肥料共分3次(移栽时、移栽后第1周以及第2周)施入盆内,3次施肥比例为2:1:1。在移栽后50d现蕾时打顶,单株留叶18片。
将赤星病菌于PDA固体培养基内在25℃环境条件中连续培养7d。待充分产生孢子后,加入10mL质量分数为1%的无菌葡萄糖溶液浸泡15min;用灭菌后的棉签在PDA培养基表面轻刮后将孢子与菌丝体从培养基内洗出,通过4层无菌纱布充分过滤除掉菌丝体,最后用1%的无菌葡萄糖溶液配制成浓度达到1.0×10个mL的烟草赤星病菌孢子悬浮液用于下一步的人工接种。
1.3试验处理
在移栽后50d,挂牌标记2个品种第3、4叶位的叶片(由下往上数,下同),使用孢子悬浮液喷雾方式开展接种(所用的孢子悬浮液于人工接种时现用现配),每个品种每个重复选择25株烟苗用于赤星病菌接种。采用小型手持喷雾器将孢子悬浮液喷酒至叶片上,使孢子悬浮液在叶面上呈现均匀分布却不下滴的程度,1%无菌葡萄糖溶液处理作为試验对照。接种后将烟株置于人工气候室中,25℃黑暗环境下保湿24h,然后开展正常的生长管理,使昼夜温度保持在27℃/23℃,相对湿度为70%。在接种后0、3、6、9、12d,每天9:3010:30采用随机方式进行取样,2个品种每个重复每次分另取样3片,将叶片去除主、侧脉并用锡箔纸包裹,利用液氮快速冷冻后置于-80℃超低温冰箱中保存,用于内源激素含量测定,重复3次。
1.4内源激素测定方法
取烟草叶片1g,加入8倍量80%预冷甲醇在4℃黑暗条件下浸泡24h。此后在4℃下18800g离心10min,收集上清液;残渣加入甲醇(体积比1:5)重复提取2次。获得的上清液合并后再次离心10min。将离心后的上清液通过C18小柱,然后依次加入1mL磷酸缓冲液(pH3.0)和乙酸乙酯,等体积萃取3次,把上层乙酸乙酯相经真空干燥后浓缩,此后加入2mL甲醇进行溶解,最后经0.45um有机系微孔滤膜过滤,将收集的样品在1100LC型高效液相色谱仪(AgilentInc,USA)上测定ABA、GA、IAA、CTK及JA含量,检测器为二极管阵列检测器(DAD)。
ET释放量参照杨暹等的方法进行测定。在接种后0、3、6、9和12d,2个处理分别取烟叶8片(第3、4叶位),立即放入500mL三角瓶中,室温下密封3h后抽气1min,用HIITACHII163型气相色谱仪测定,每个处理重复3次,每一重复抽气3针。气相色谱参数设定为:火焰离子化检测器FD色谱柱温90℃,色谱进样器温度100℃,载气是N2,流速25mL/min利用纯乙烯绘制标准曲线样品的色谱峰值经过换算得到ET释放量。ET释放量=乙烯浓度(三角瓶体积烟草叶片体积(密封时间×烟草叶片重量)×0.8×储温。
1.5数据分析
使用SPSS17.0软件与MicrosoftExcel2013对所测定的数据进行统计分析和作图,采用Duncans新复极差检测法进行数据显著性分析。
2结果
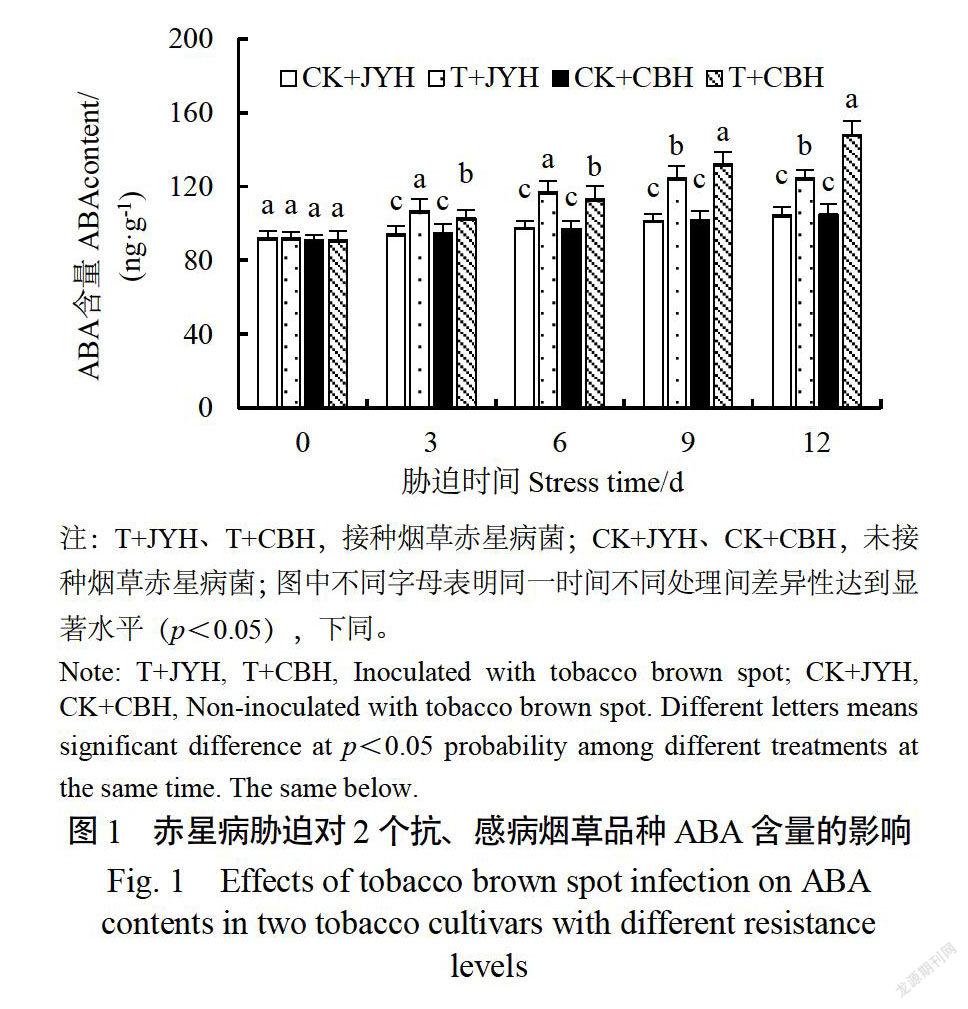
2.1赤星病胁迫对抗、感病烟草品种ABA含量的影响
正常生长条件下,2个品种的ABA含量随烟株生长发育均上升(图1)。与对照相比,赤星病胁迫致使抗、感病品种的ABA含量均显著增加(p《0.05)。在胁迫3、6d时,JYH的ABA含量分是CBH的1.04、1.05倍,达到显著差异水平(p《0.05)。此后,CBH的ABA含量大幅度上升;在胁迫9、12d时,CBH的ABA含量分别是JYH的1.06、1.11倍,差异也达到显著水平(p《0.05)。
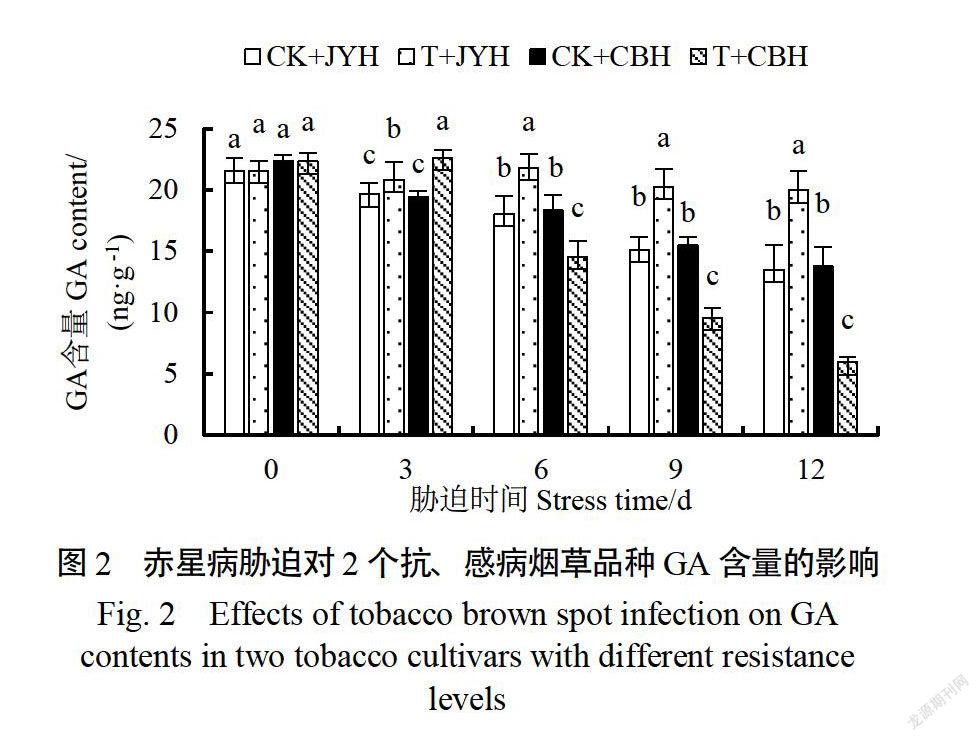
2.2赤星病胁迫对抗、感病烟草品种GA含量的影响
正常环境中,随着烟株生长发育,2个品种的GA含量均下降,但降低幅度较小(图2)。赤星病胁迫使JYH的GA含量显著高于对照(p《0.05),而CBH的GA含量表现出先缓慢增加后急剧下降的变化趋势,且在处理3d时达到最大值,为对照的1.16倍,JYH的1.09倍,均达到显著差异水平(p《005)。在赤星病胁迫6-12d时,JYH的GA含量始终显著高于CHB(p《0.05),分为CBH的1.50、2.12和3.36倍。
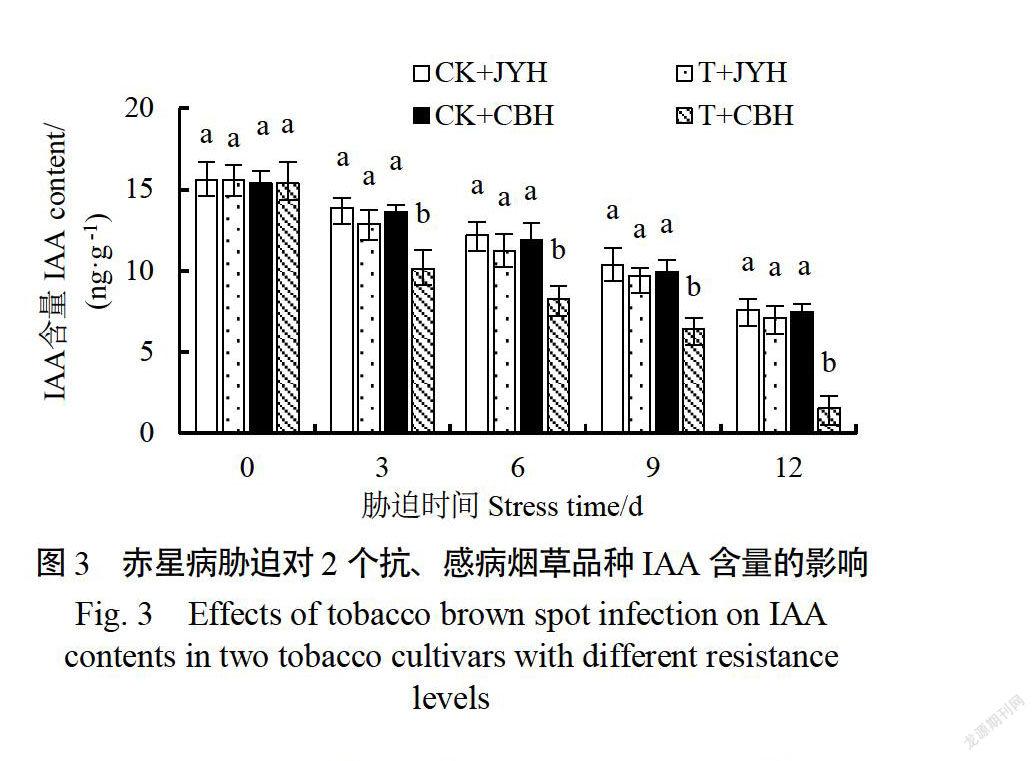
2.3赤星病胁迫对抗、感病烟草品种IAA含量的影响
如图3所示,在接种赤星病菌后,不同抗性品种的IAA含量均出现降低。胁迫12d时,与对照相比,JYH、CBH的IAA含量降幅分别为7.09%、79.44%在胁迫3~12d时,JYH的IAA含量始终显著高于CBH(《0.05),分别是CBH的0.98、1.36、1.50和4.60倍。
2.4赤星病胁迫对抗、感病烟草品种CK含量的影响
正常条件下,不同抗性品种的CTK含量均呈缓慢下降趋势(图4)。在胁迫3-9d时,JYH的CTK含量与对照无显著差异;在胁迫12d,降幅达到22.79%,与对照相比差异显著(《0.05)。赤星病胁迫使CBH的CTK含量急剧下降,显著低于对照(p《0.05)。在胁迫3-12d时,JYH的CTK含量分别为CBH的137、1.47、1.67和1.74倍,且均呈显著差异水平(p《0.05)。
2.5赤星病胁迫对抗、感病烟草品种ET释放量的影响
图5显示,正常生长发育环境下,2个品种的ET释放量呈缓慢上升趋势。赤星病胁迫造成抗、感病品种的ET释放量表现出较大的上升幅度。在赤星病胁迫处理0-6d时,JYH的ET释放量小幅上升,但与对照差异不显著;胁迫9、12d时,JYH的ET释放量显著高于对照(p《0.05)。CBH的ET释放量在胁迫处理612d时显著高于对照与JYH(p《0.05),分别是JYH的1.23、15和1.08倍。
2.6赤星病胁迫对抗、感病烟草品种JA含量的影响
如图6所示,在正常生长条件下,抗、感病品种的JA含量逐渐降低。在赤星病胁迫初期JYH的JA含量增加,在胁迫3d时显著高于对照(p《0.05)之后与对照无显著差异。在胁迫312d,JYH的JA含量始终显著高于CBH(p《0.05),分别为CBH的1.49、1.46、1.96和2.48倍;而此时CBH的JA含量呈现持续下降趋势,且显著低于对照(p《0.05)。
3讨论
内源激素是植物抵御病原菌侵染的有效防线。本试验结果显示,在赤星病胁迫下,净叶黄、长脖黄的ABA含量和ET释放量均出現上升,但GA、IAA、CTK则呈降低趋势;与净叶黄相比,长脖黄的ABA含量增幅较小,而GA、IAA、CIK与JA含量降幅较大,并且ET释放量较高。这与钟丽娟等田、吴俊江等的研究结果基本一致。本文研究充分表明不同抗性品种所含的内源激素在赤星病菌侵染时出现不同变化,从而对抗病性发挥作用。周青的研究结果证实,植物保卫细胞中的ABA能够在逆境条件下对外排放,造成气孔关闭;同时促进脯氨酸与乙烯的大量合成以维持植株内的水分平衡,最终提高植物抗性。ABA不仅能够促进植物的光合作用与光合产物运输,还可以提高根系的吸水速率,有助于向地上部供水。在本研究中,净叶黄含有的ABA含量在赤星病胁迫3-6d显著高于长脖黄,可维持叶片细胞膜的稳定性,保证光合反应与光合活动生成物转运的正常进行,强其抗赤星病能力。但ABA长时间的过量增加会导致植物叶片加快脱落,造成植株提前老化死亡在赤星病胁迫处理的9~12d,感病品种长脖黄的ABA积累量显著高于净叶黄,并且长脖黄的ET释放量在胁迫612d也明显大于净叶黄,导致其开始出现赤星病病症,从而加快衰老死亡。感病品种在赤星病胁迫后期具有较大的ABA积累量、ET释放量,显示出其细胞膜结构被严重破坏,导致叶片的衰老死亡活动出现大大提前。这与文献[12-13]的研究结论基本一致。
长脖黄中的GA含量在赤星病胁迫下表现出先增加再降低的动态变化趋势,而净叶黄的GA含量在胁迫处理下显著高于对照,显示出早期的赤星病胁迫有助于不同抗性品种加快合成GA。但伴随胁迫处理时间的推移,这类促进效应在感病品种中明显降低,其GA调节性能受到明显抑制,而最终表现为患病区域坏死成病斑。叶黄的GA含量在胁迫612d均显著高于长脖黄,表明抗病品种却能够通过增加GA含量减轻不良影响,保证叶片的正常生长发育,缓解赤星病胁迫的抑制效应。蔡刘体的研究结果显示,小麦在感染白粉病初期GA含量迅速上升,之后下降并维持在较高水平,这与本研究结果相接近。
具有抗萎病特性的榆树品种能够在受到侵染后比感病品种更早产生导管侵填体物质,这和具有较高含量的IAA从而有助于出现抗病障碍存在紧密联系。CTK有利于促进物质运输,对阻止叶绿体降解、维持细胞膜结构的完整性、推迟衰老死亡和清除活性氧自由基(ROS)破坏等活动均发挥明显的积极影响,其含量出现降低是造成植物叶片的气孔关闭、光合活动受抑制的一个重要因素KOVAC等研究发现,在受马铃薯YNN病毒侵染之后,JA含量表现出瞬时大量激,最终诱导植物对病毒的抗病能力。在赤星病胁迫环境中,抗病品种净叶黄所含的IAA、CTK和JA含量均显著高于感病品种长脖黄,使其不会因CTK含量快速降低而导致膜脂过氧化水平加剧与衰老死亡活动加速,而较高含量的IAA可易于生成抗病障碍,加之JA含量的上升可增强一系列抗氧化防御酶及与抗坏血酸-谷胱甘肽循环相关的基因表达,继而提升ROS清除能力,有助于保持叶片细胞结构的完整程度,最终提高抗赤星病能力。
4结论
本研究发现,在赤星病胁迫下,抗病烟草品种中的内源激素积极参与对赤星病菌侵染的防御反应,ABA和JA幅较大,GA、IAA、CTK下降缓慢,ET释放量较少,从而能够保持烟株的代谢平衡和稳定性,在一定程度上抑制或者减轻赤星病胁迫的不利影响;但感病品种的ABA含量在赤星病胁迫下的增幅较小,GA、IAA、CTK和JA下降迅速,ET释放量较高,最终呈现为感病而加快衰老死亡。内源激素的含量差异与变化幅度可以反映不同抗性烟草品种抗赤星病能力。
参考文献
[1]旷永洁,柳浪,严芳,等.水稻与病原物互作中植物激素功能的研究进展.生物技术通报,2018,34(2):74-86.
[2]邹华松,柔伟,吴薇.柑橘溃疡病抗感病性机制研究进展[J].森林与环境学报,2018,38(2):234239
[3]徐以华.水稻穗枯病菌的分离及品种内源激素与抗病性关系研究[D]南宁:广西大学,2018
[4]钟丽娟,赵秀香,贾玉才,等.烟草感染马铃薯Y病毒脉坏死株系后内源激素的变化安徽农业科学,2006,34(15):3724-3725
[5]吴俊江,刘丽君,高明杰,等不同抗性大豆品种(系)接种灰斑病1、7号生理小种后内源激素变化规律的研究[J].大豆科学2001,20(1):14-17
[6]童治军,张谊寒,陈学军,等.雪茄烟品种Beinharti000赤星病抗性基因的QTL定位.作物学报,2019,45(3):47482
[7]杨铁钊.烟草育种学[M].第二版.北京:中国农业出版社,2011:285
[8]杨暹,陈晓燕.不同氮营养下炭疽病菌侵染对菜心叶片内源激素的影啊.应用生态学报,2005,16(5):919923
[9]周青.逆境胁迫下植物积累脱落酸的生理生态学意义.农业现代化研究,1993,14(4):231-235
[10] PARIDA A K, DAS A B Salt tolerance and salinity effects on plants a reviewj]. Ecotoxicology and Environmental Safety, 2005, 60(3)324-349.
[11]张明生,谢波,谈锋.水分胁迫下甘薯内源激素的变化与品种抗旱性的关系,中国农业科学,2002,35(5):489-501
[12]宋凤鸣,郑重.乙烯产生的抑制与氟乐灵诱导棉苗对枯萎病的抗性有关.植物生理学报,1998,24(2):11-118
[13] COOPER W, BOUZAYEN M, HAMILTON A, et al. Use of transgenic plants to study the role of ethylene and polygalacturonase during infection of tomato fruit by Colletotrichum gloeosporioides [J]Plant Pathology, 1998, 47(3): 308-316
[14]蔡刘体玉米索核苷单克隆抗体的制备及小麦与白粉病互作初期激素作用的分析[D]南京:南京农业大学,2003
[15] OUELLETTE G B, RIOUX D Anatomical and physiological aspects of resistance to dutch elm disease A]/BLANCHETTE R A, BIGGS A R. Defense mechanisms of woody plants against fungi[M]Heidelberg: Springer Heidelberg, 1992: 257-307
[16]汪寶卿,慈敦伟,张礼风,等.同化物供应和内源激素信号对大豆花发育的调控.大豆科学,2010,29(5):878-882
[17] AHMED S, NAWATA E, SAKURATANI T. Changes of endogenous ABA and ACC, and their correlations to photosynthesis and water relations in mungbean (Vigna radiate (L)Wilczak cv KPS1)during waterlogging[J].Environmental and Experimental Botany,2006,57(3):278-284.
[8] KOVAC M, MULLERA, JARH D M, et al. Multiple hormone analysis indicated involvement of jasmonate signaling in the early defence of potato to potato virus YNTN[J]. Biologia Plantarum, 200953(1):195-199.
[19] KUMAR GJ, REDDY A M, NAIK S T, et al. Jasmonic acid induced changes in protein pattern, antioxidative enzyme activities and peroxidase Isozymes In peanut seedlings [J] Biologia Plantarum,2006,50(2):219-226
[20] SASAKI-SEKIMOTO Y, TAKI N, OBAYASHI T, et al Coordinated activation of metabolic pathways for antioxidants and defence compounds by jasmonates and their roles in stress tolerance in Arabidopsis [J]. Plant Journal, 2005, 44(4): 653-668
猜你喜欢
新农业(2021年21期)2021-11-20
人物画报(2020年22期)2020-03-09
长江蔬菜·学术版(2016年12期)2017-01-12
湖北农业科学(2016年18期)2016-12-08
股市动态分析(2016年6期)2016-09-29
商界评论(2016年4期)2016-04-20
长江蔬菜·学术版(2015年1期)2015-07-10
绿色中国(2015年6期)2015-07-03
新民周刊(2014年22期)2014-06-24
农家顾问(2009年5期)2009-05-27
