
水稻高节位分蘖的形态特征及遗传行为
2021-05-20 09:45霍川王世全沈俊宏曾鸿燕
中国农业科技导报 2021年5期
霍川, 王世全, 沈俊宏, 曾鸿燕
(1.四川农业大学水稻研究所, 四川 温江 611130; 2.四川省资阳市农业技术推广站, 四川 资阳 641400)
水稻是我国重要的粮食作物之一,常年种植面积约占全国粮食作物总面积的30%,产量接近粮食总产量的三分之一[1]。近20年来,我国水稻种植面积一直稳定在3 000万hm2左右,与20世纪70年代比较种植面积有所下降,但科学技术的进步促进单产水平不断提高,总产量仍然稳定在2亿t以上[2]。尽管我国水稻种植面积居世界第二,总产量居世界第一,但单产水平平均不足7 000 kg·hm-2,仅居全世界第13位(2013年)[2]。近年来,随着人口的增加和人民对美好生活的向往,加上工业用量增多,我国水稻进口量呈上升趋势。所以提高水稻产量和品质是水稻产业发展和保障国家粮食安全的主要任务[3]。
水稻产量是由单位面积的穗数、每穗粒数和千粒重三个基本因素构成的,而单位面积的穗数又是由株数、单株分蘖数、分蘖成穗率三者组成的。当单位面积株数一定时,单株分蘖数和分蘖成穗率便成为穗数的重要决定因素[4],分蘖数通过影响穗数对单位面积产量提高产生至关重要的作用。研究水稻分蘖的表型特性及其遗传行为,可为创制分蘖较多的育种材料或品种奠定较好的理论基础,为水稻的高产育种提供重要的指导价值。水稻分蘖能否发生、分蘖多少、分蘖节位高低既有遗传原因[5-8],也有环境的影响[9-19]。本课题组在野生型水稻恢复系R818中发现并筛选出一个高节位分蘖突变体W33,栽种过程中发现,该突变体高节位分蘖与高分蘖成穗率性状表现稳定。本研究拟以W33为试验材料,研究水稻高节位分蘖的表型特性与遗传行为,为进一步开展水稻高节位分蘖理论研究与利用途径提供参考和试验方法。
1 材料和方法
1.1 试验材料
以水稻高节位分蘖突变体W33和水稻野生型品系R818为试验材料,来源于四川农业大学水稻研究所。其中W33是在水稻恢复系R818中发现并筛选出的一个高节位分蘖且分蘖成穗率较高的突变体,该突变体植株较矮、抗倒性强,除穗颈节外每节均会分蘖成穗,分蘖数多、成穗率高,穗小、穗粒数少,但籽粒形状、大小、千粒重等与野生型没有明显差异。R818即蜀恢818,为1个选育的三系恢复系。
1.2 试验设计与方法
分蘖形态特性观察采用水培(幼苗期)和盆栽(分蘖期)试验,表型遗传行为研究采用田间试验。
幼苗期水培试验:将W33和R818种子用3%双氧水浸泡24 h以消毒和打破休眠,用蒸馏水洗净后放入38 ℃恒温培养箱中黑暗条件下催芽至露白,将露白发芽的种子转移至下部镂空的96孔PCR板,再将PCR板放置于pH 5.5的水稻营养液中,然后在PRX-800B型人工气候培养箱(上海颐习设备有限公司)中培养7 d,期间更换营养液一次。培养箱条件:31 ℃/24 ℃(白天/夜晚);光照时间7:30—19:30;光照强度3 000 lx。水培液配方采用国际水稻所配方标准[20]。
盆栽试验:于2017年3月28日,在四川农业大学温江校区校园内开展,花盆选择高40 cm,Φ30 cm的圆柱形塑料钵,土壤用紫色砂壤土,每钵用N15∶P15∶K15三元复合肥,施用量为94.4 kg·hm-2。2017年4月10日,每个塑料钵内分别播种催芽的R818或W33种子6粒。钵内种子均播成三角形。待种子发芽长成1叶1心时,间苗每盆保留3棵,4叶1心时每钵施用N15∶P15∶K15三元复合肥和尿素各47.2 kg·hm-2,保持淹水3~5 cm至新种子成熟。治虫、防病、除草等措施同大田生产。
田间试验:于2016年春季和秋季,分别在四川农业大学温江校区实验农场和海南省陵水县四川农业大学海南南繁基地,将W33和R818分别进行两地各一次种植自交纯化,并重点观察W33高节位分蘖和R818分蘖特性是否存在环境效应变化。2016年冬季,取自交纯化后的R818和W33种子各50粒,播种在海南省陵水县四川农业大学海南南繁基地,开花期进行正交(R818×W33)和反交(W33×R818)获得正反交F1杂交种子。2017年春季,取部分正反交F1杂交种子和自交纯化后的W33、R818种子,种植于四川农业大学温江校区实验农场,观察W33高节位分蘖特性是否存在典型显隐性遗传关系和细胞质遗传特性,同时将W33、R818各自分别与部分正反交F1杂合体进行回交获得回交种子,另一部分正反交F1杂交种在抽穗时分别进行套袋自交获得F2种子。2018年春季,在四川农业大学温江校区实验农场,将自交纯化后的W33、R818、正反交F1、正反交F1与W33的回交后代、正反交F1与R818的回交后代、正反交F2种子种于田间,其中W33、R818、正反交F1每个种植2行40株,回交后代各种植12行240株,F2各种植15行300株。
所有田间试验种植规格均为行距0.25 m、窝距0.2 m,单株种植,折合种植密度为200 010株·hm-2,田间施肥、灌水、治虫、防病、除草等措施同当地大田生产。
1.3 指标检测及方法
将水培7 d的W33和野生型R818幼苗各取10株,观察单株形态特征,并测量幼苗高度、主根长度;将幼苗下部浸入水中,观察并记录不定根条数,测量不定根长度。
记载盆栽试验参试材料生长发育的生物学时期,包括分蘖期、拔节期、孕穗期、抽穗期和成熟期。在分蘖期和拔节期观察参试材料的植株形态特征、分蘖动态及形态。分别从R1~R3和W1~W3号钵中依次取三次样,每次取1株测量植株高度、记载主茎叶片数、主茎叶长度、主茎分蘖数、次生分蘖数。在孕穗期,从R4~R13和W4~W13号钵中选取主茎叶片数相同的W33和R818各10个单株,从开始孕穗第5 d进行取样,用手术刀片分别对W33和R818倒1节、倒2节和倒3节进行环剥(注意不能伤及分蘖芽),取下腋芽测量长度,并迅速用卡诺固定液进行固定保存,在接下来的45 d内每隔5 d取一次样,每次取1株,重复操作。在抽穗期,分别从R14和W14号钵中取R818和W33发育正常、基本成熟尚未开放的主茎颖花各3朵,用镊子取1个花药,置于载玻片,用镊子将花药捣碎,滴加1~2滴I2-KIStain,盖上盖玻片,每个材料制成3张片子,立即放在1 500倍解剖显微镜下观察,每片取5~6个视野,统计每个视野花粉的染色率。分别从R14和W14号钵中取R818和W33全部植株测量株高和主茎叶片长度,解剖统计最终分蘖数,包括一次分蘖、二次分蘖、三次分蘖、四次分蘖。籽粒灌浆成熟后,分别从R15和W15号钵中取出R818和W33全部植株,测量主茎和分蘖稻穗的穗长,统计有效分蘖数、稻穗一次和二次枝梗数、颖花数、结实粒数(穗粒数)、结实率;分单穗脱粒晒干后测定单穗粒重、单株粒重和千粒重。
记载田间试验各参试材料的生物学时期,在抽穗期调查记载W33是否存在高节位分离现象,杂交F1是否有高节位分蘖出现,以及正反交F1高节位分蘖现象是否有差异,分离世代有高节位分蘖和无高节位分蘖性状植株数。
1.4 数据统计与分析
用Microsoft Excel 2003进行试验数据处理,用SPSS 19.0计算平均数、进行显著性分析和2遗传适合性检验。
2 结果与分析
2.1 幼苗形态特征
水培7 d后的水稻幼苗特征结果(表1)显示,W33幼苗的株高、主根长度、不定根条数、不定根长均与R818无显著差异(P>0.05)。表明W33幼苗的表型特征和野生型R818并无显著差异,它们的地上部分和地下部分均能生长正常。

表1 水培下R818与W33的幼苗特征
2.2 分蘖特性
从图1可以看出,在分蘖前期,R818的可见叶为7叶,而W33的可见叶为8叶,二者相差1片可见叶。但二者的生长速度、植株高度、叶长、主茎分蘖数等无显著差异。W33和R818的主茎上均可见4个一次分蘖,但W33主茎的第一和第二叶位上已经出现了明显的二次分蘖,而R818未见二次分蘖出现。

注:红色箭头指二次分蘖
进入拔节期后,W33的植株高度平均为(55.3±3.12) cm(表2),比R818矮21.6 cm,而单株平均二次分蘖数为(4.60±0.70) 个,平均三次分蘖数为(3.30±0.56) 个,分别比R818多2.3个,差异均达极显著水平(P<0.01);主茎叶片平均长度(30.2±2.24) cm,比R818短6.30 cm,差异显著(P<0.05)。但W33的主茎平均可见叶片数(12.3±0.62)和平均一次分蘖数(7.80±0.01)与R818比较均无显著差异(P>0.05)。表明进入拔节期后,W33快速分蘖的优势已逐渐显现出来,但主茎叶片长度显著短于R818,株高极显著低于R818。
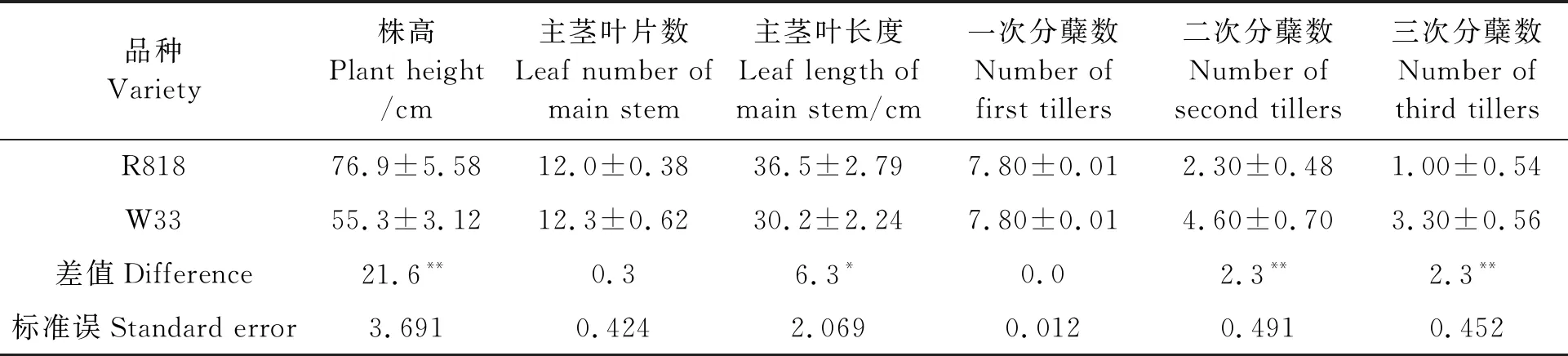
表2 拔节期R818与W33的形态特征
孕穗期两个参试水稻材料的腋芽生长状态结果见图2,可以看出,在孕穗期间,W33的腋芽生长迅速,倒一节、倒二节和倒三节腋芽几乎处于同步生长状态;而R818的倒一节和倒二节腋芽基本处于休眠状态,倒三节腋芽生长速度缓慢。

图2 孕穗期R818与W33的茎倒一至倒三节环剥图
环剥后的主茎倒二节和倒三节的腋芽长度测量结果见图3。可以看出,孕穗15 d时,W33的倒二节腋芽长度为12 cm左右,比R818长约12 cm;倒三节腋芽长度为19 cm左右,比R818长约18 cm,二者同期差异极显著(|t|=3.681>t0.01,9=3.250)。孕穗15 d后,W33的倒二节和倒三节腋芽仍保持直线生长状态至孕穗后25 d,此后仍在缓慢伸长,且倒二节腋芽与倒三节腋芽生长速度基本同步;而R818的倒二节腋芽几乎完全进入休眠状态,倒三节腋芽生长速度十分缓慢至孕穗25 d基本停止。表明进入孕穗期后W33腋芽特别是高节位腋芽仍处于活跃、快速生长状态,而R818高节位腋芽逐渐进入休眠状态。
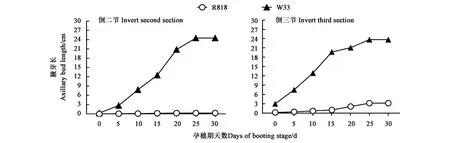
图3 孕穗期倒二节和倒三节的腋芽生长
抽穗期的R818与W33的植株及分蘖特性结果(表3)显示,进入抽穗期,W33的平均株高为71.3 cm,比R818矮32.2 cm,差异极显著(P<0.01); W33的快速分蘖优势再次凸显,平均一次分蘖数、二次分蘖数、三次分蘖数、四次分蘖数分别达到(16.4±2.9)(26.9±7.9)(20.8±6.6)和(15.6±5.3) 个,均极显著高于R818(P<0.01),单株平均分蘖总数达到79.7个,是R818的7.5倍;但主茎叶均长只有(30.0±2.2) cm,比R818短5.8 cm,差异显著(P<0.05)。

表3 抽穗期R818与W33的植株及分蘖特征
当R818的主茎处于抽穗期时,W33的主茎还处于孕穗期,根据生育期统计,W33的抽穗期平均比R818晚15 d左右。取孕穗期的R818和W33正常颖花的花粉粒进行碘染观察,W33的碘染率为98.3%,R818的碘染率为98.7%,两者正常颖花的花粉粒碘染结果没有显著差异(P>0.05)。表明W33的主茎生殖发育进程可能会因其旺盛的高节位分蘖特性有延缓的趋势,但并不影响其雄性生殖细胞的活性。
2.3 穗部特征
将盆栽条件下的W33和R818植株叶片去除后,露出全部穗子,结果(表4)发现,W33的平均有效穗数为42.80,而R818的平均有效穗数为6.20,约为W33的1/7。R818低节位的一次分蘖上仅有一个穗子,几乎没有二次分蘖成穗的情况;而W33低节位的一次分蘖上至少有两个以上的穗子,即一次分蘖上长出的二次甚至三次分蘖均能成穗,平均成穗率为81.5%,与R818一次分蘖成穗率(82.1%)接近。
部分穗部性状统计分析结果(表4)表明,W33的单株平均有效穗数显著高于R818,而其单个稻穗的一次枝梗数、二次枝梗数、穗长、单穗粒数和平均单穗产量均显著低于R818(P<0.05);但结实率、千粒重和平均单株产量与R818无显著差异。W33的单株产量略高于R818,可能是因为W33分蘖优势明显,以致有效穗数显著增多。
2.4 表型遗传特征
W33和R818植株在成都温江、海南陵水两地各种植1次,发现突变体W33的高节位分蘖性状不因生态环境或栽培措施的改变而改变,所有单株均存在高节位分蘖现象,单株分蘖数68.8~79.3个,平均72.6个(数据未列出);野生型R818在两种生态环境条件下均只存在基部或低位分蘖现象,单株分蘖数6.1~9.3个,平均7.7个。表明突变体W33的高节位分蘖性状未出现遗传分离现象,该性状遗传表现稳定,R818无高节位分蘖特性。用2次自交纯化后的W33与野生型R818组配的正反交F1代均未表现为高位分蘖现象,说明高节位分蘖性状属于细胞核基因控制性状,且表现为隐性遗传,与细胞质遗传无关。

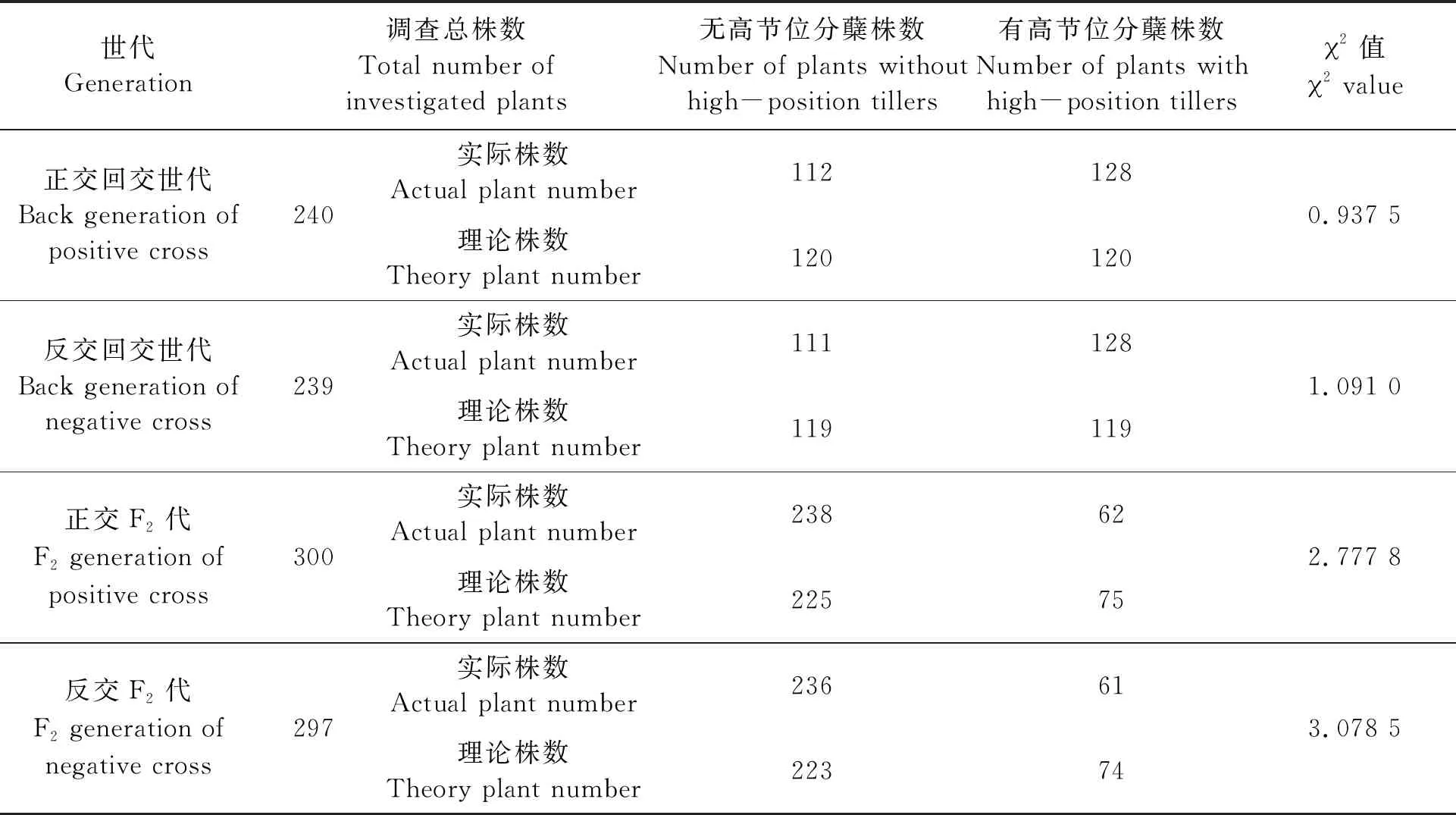
表5 不同世代分离群体的高节位分蘖统计
3 讨论
3.1 W33具有典型的高节位分蘖特性
分蘖是水稻等禾本科植物经过长期进化发育保留下来的重要生物学和形态学特征。一般认为,水稻分蘖主要发生在植株基部密集的节位上[21],但也有研究和生产实践证明,环境条件改变或农事操作不当,如水温、环境温度和土壤温度改变[10,17,19]、淹水太深[4]和密度过大[4,10]、秧龄过长[18]、分蘖前期水分和氮、磷、钾等养分供应不足[11-15]、农药使用不当[22]等均可能诱发高节位分蘖。莫永[22]研究发现,一些分蘖力强的品种也常出现高节位分蘖,且高节位分蘖在籼稻品种出现机率大于粳稻和糯稻,多穗型品种出现机率大于大穗型品种,杂交水稻出现机率大于常规水稻,野生稻更易出现高节位分蘖。这些研究的共同特点是以野生型水稻品种为试验材料,通过环境条件或栽培措施的改变来影响分蘖数量和分蘖节位的变化,并发现水稻能否出现分蘖、分蘖多少、分蘖节位高低,是基因型与环境互作的最终结果。本研究发现,突变体W33在其生长发育过程中,幼苗期间地上和地下表型特征和其野生型并无差异,但进入分蘖期后,多分蘖和高节位分蘖优势逐渐显现出来,拔节期多分蘖和高节位分蘖优势更加明显,到孕穗后期其腋芽特别是高节位腋芽仍处于活跃快速生长阶段(除穗颈节外每节均会分蘖),最终导致分蘖数猛增,这是突变体W33与野生型R818在分蘖特性上的最大区别特征。其次,W33穗子较小、单穗穗粒数较少是区别于野生型R818的明显穗部形态特征,但W33分蘖数多、成穗率高,籽粒形状、大小、千粒重与R818没有明显差异,其单株产量最终能达到与野生型R818的相当水平。W33植株相对较矮、叶片平均长度相对较短,可能是因为W33植株在增加分蘖过程中,将更多的营养物质用于分蘖建成及生长发育,从而限制了株高和叶长的增加。
3.2 高节位分蘖特性表现为隐性遗传
关于水稻多分蘖性状的遗传已有一些研究报道,段远霖等[23]以1个受一对隐性基因控制的多分蘖突变体det1为材料,将该隐性突变基因定位在6号染色体的长臂端;江海湃等[24]将一对隐性基因控制的多分蘖基因HDT1-2定位在4号染色体上;薛晶晶等[25]将一对隐性单基因控制的多分蘖矮秆基因D63定位于8号染色体短臂端;张艳培等[26]以1株受1对隐性基因控制的多蘖突变体f2-132为材料,将该隐性基因定位在4号染色体上。这些研究均证明水稻多分蘖性状受隐性基因控制,在表型遗传行为上符合孟德尔遗传分离定律。但王永胜等[6]从栽培稻粳籼89种子EMS化学诱变M2代植株中筛选到1株受两对显性等位基因控制的多分蘖突变体ext370,李万昌等[27]在新稻18中发现1株多分蘖显性突变体植株HT1,这些研究均未涉及高节位分蘖问题。谈雅静[28]对一个高节位分蘖的小剑叶突变体sfl的表型及农艺性状进行研究发现,突变体sfl较野生型穗期推迟,剑叶变小变短,剑叶叶舌消失,穗变小,穗杆变短,遗传分析发现sfl突变体为单基因隐性突变。本研究表明,高节位分蘖突变体W33的高节位多分蘖性状属于细胞核基因控制性状,在W33与其野生型水稻R818的正反交F1与W33的回交后代中,无高节位分蘖和有高节位分蘖的分离符合1∶1分离比;在正反交F2世代中,无高节位分蘖和有高节位分蘖的分离符合3∶1分离比。表明本研究所用突变体W33高节位分蘖性状的遗传受一对隐性核基因控制,W33是该基因的隐性突变体。这一结果与已有的研究结果[23-26]类似,与谈雅静[28]研究的突变体sfl的有高节位分蘖和无高节位分蘖在回交世代及自交世代的分离比结果一致。然而,该基因所在的染色体位点还有待通过基因定位方法进行锁定。
3.3 高节位分蘖特性具有一定的利用价值
众所周知,单位面积穗数、每穗粒数和千粒重是构成水稻稻谷产量的三要素,只有各个要素协调增长,使单位面积总粒数达到最多,千粒重相对稳定或提高的情况下才能获得高产。张恒栋等[4]研究认为,当单位面积基本株数一定时,单株分蘖数和分蘖成穗率便是每亩穗数的重要决定因素;孙长占等[29]研究表明,单位面积穴数,每穴穗数和每穗实粒数基本上决定了水稻产量的大部分变异,它们对产量形成的相对重要性依次为:每穴有效穗数>单位面积穴数>每穗实粒数。这些研究均证明单位面积穗数(由基本苗、分蘖数和分蘖成穗率决定)是影响水稻产量的重要因素。本研究表明尽管高节位多分蘖突变体W33的单穗穗短,实粒数少,产量较低,但其旺盛的分蘖力和较高的分蘖成穗率优势能弥补其单穗产量较低的缺陷,最终达到与野生型R818单株产量相当的水平。因此,本研究认为在穗数型水稻品种选育方面,高节位分蘖性状对于增强水稻分蘖力,增加单位面积穗数,或许具有一定的利用价值。
猜你喜欢
安徽农业科学(2022年19期)2022-10-29
安徽农业大学学报(2022年3期)2022-10-25
中国烟草科学(2022年2期)2022-05-27
植物研究(2022年2期)2022-03-02
塔里木大学学报(2021年1期)2021-03-31
南方农业·上旬(2021年1期)2021-03-02
中国烟草科学(2019年1期)2019-09-10
吉林农业(2017年1期)2017-02-16
江苏农业科学(2016年6期)2016-07-25
江苏农业科学(2016年6期)2016-07-25
