
七种黄精属植物的光合特性
2021-07-09 03:09马英姿周诗雨
经济林研究 2021年2期
刘 熙,宋 荣,马英姿,周诗雨,刘 智
(1.中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;2.湖南省农业环境生态研究所,湖南 长沙 410125)
黄精为百合科Liliaceae 黄精属Polygonatum[1]。多年生草本植物的总称,中国约有31 种,在国内分布较广泛,该属植物有肉质根状茎,形似于圆柱形、结节处有时不规则,一般生长在灌丛、林下和阴凉处,具有保健、营养、滋补、治疗肾虚、精血不足、肺虚、干咳等药用功效,同时在抗衰老、调节自身免疫、降低机体血脂、增强机体记忆反应能力、抗恶性肿瘤、抗菌等方面也具有潜在的生物医学价值[2-3]。目前,黄精种苗资源在很大程度上依赖于野生黄精种质资源,随着现代社会对黄精的需求日益增加,我国黄精野生资源面临枯竭和生态环境严重破坏的风险,林下栽培黄精可以减少黄精野生资源的消耗,并满足市场对黄精的需求,同时,选育出优良的黄精种质资源已成为亟待解决的关键问题[4]。
农作物的产量取决于光合作用过程中光合产物的积累和分布,如何高效地利用太阳能增强植物的光合作用,目前仍然是现代农业发展中的一个根本性问题。选育高光效种质就是一种提高作物产量和品质的有效方法,选择高光效种质的重要指标是其光合特性[5-6]。植物的光合特性主要包括光合作用日变化、光合速率、光饱和点和光补偿点[7]。光合作用的日变化曲线可以直接反映各种植物之间的内在节律差异,还能够反映各种植物之间环境适应性的差异,因此可以代表各种植物的生理特性和其它环境因素对它们的综合影响[8-9]。本研究对湖南省内主要种植的多花黄精P.cyrtonema、黄精P.sibiricum、滇黄精P.kingianum、点花黄精P.punctatum、湖北黄精P.zanlanscianense、卷叶黄精P.cirrhifolium、长梗黄精P.filipes的光合特性进行了系统的测定,旨在对黄精属植物的生理生态习性有更为精准的掌握,探讨其对环境适应能力的差异,为选择黄精属植物的优良种质和科学高效地栽培利用提供理论依据。
1 材料与方法
1.1 试验地与试验材料
试验地位于湖南省长沙市农业环境生态研究所基地。属亚热带季风性湿润气候,年平均气温为17.5 ℃,年平均最高气温为21 ℃,年平均最低气温为14 ℃[10]。
本试验材料均采自湖南省安化县及湖南省农科院的长沙县高桥基地3年生黄精属植物栽培种,于2019年4月黄精出苗期整株采集移植。7 种黄精属植物植株均经过湖南中医药大学王智老师鉴定,分别为点花黄精、多花黄精、滇黄精、湖北黄精、黄精、卷叶黄精和长梗黄精,栽培于林下环境中。
1.2 方法
光合指标测定时间为2020年7月中旬。测定时黄精属植物均为4年生植株结实期,连续数日选择晴朗天气测定其光响应曲线、CO2响应曲线和日变化,每种黄精属植物测定3 株,每株测定3片叶,选择中上部位同一水平、成熟无病虫害的叶片进行测定。
1.2.1 叶片叶绿素含量的测定
叶绿素含量用丙酮-乙醇混合法测定,丙酮∶乙醇(无水)=5∶5。取新鲜叶片0.1 g,切碎,放入试管中,加入25 mL 溶液,密封,在暗处放置6~8 h,然后在470、663、645 nm 进行比色,得到A值,根据Arnon法公式计算各提取物中叶绿素含量,重复测定3 次,计算其平均值[11-12]。
1.2.2 光合速率相关指标测定
以便携式光合作用测试仪Li-6400 为测量仪器,于晴天测定日变化,共测量7 次,时间段分别 为7:00、9:00、11:00、13:00、15:00、17:00、18:00。每个种随机选取3 株植物,每株选取3 片中上部位同一水平、成熟无病虫害的叶片进行测定。测定7 种黄精属植物叶片的净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)、光合有效辐射(PAR)、相对湿度(RH)和大气温度(Tair),水分利用效率(WUE)=Pn/Tr,对每片叶测定3 次,测定数据取平均值。
1.2.3 光响应曲线的测定
选取7 种黄精属植物中间部位生长状况较好的成熟叶片,在晴天用Li-6400 测定其光响应曲线。仪器叶片室采用红、蓝光光源,光响应曲线测量参数设定如下:参比室内CO2浓度约为400 μmol·mol-1,叶室温度范围控制在(30±0.5) ℃,相对湿度范围控制在(40±5)%,光响应曲线的光合有效辐射强度设定在2 000、1 800、1 500、1 200、1 000、800、600、400、200、100、50、0 μmol·m-2·s-1共12 个光照强度梯度。仪器开机时,预先设定估计的饱和光强和最佳CO2浓度,光诱导黄精叶片30 min,等到仪器测得的净光合速率(Pn)参数稳定在小数点后1 位数时记录数据。7 种黄精属植物均随机选定3 株生长良好的植株,每株选择3 片中上部位同一水平、成熟无病虫害的叶片,尽量保持叶片在植株上同一位置测量。后续采取叶子飘直角双曲线修正模型拟合光响应曲线,得到饱和光强(Isat)、初始斜率(α)、最大净光合速率(Pnmax)、暗呼吸速率(Rd)和光补偿点(Ic)等光合参数[13-14]。
1.2.4 CO2 响应曲线的测定
选取7 种黄精属植物中部生长条件较好的成熟叶片,在晴天用Li-6400 测定其CO2响应曲线。该仪器配有CO2注射器,可为叶片室提供稳定可调的CO2气源,测定中使用CO2小钢瓶提供测定所需要输出的稳定CO2浓度。在测定过程中,叶室内的光合有效辐射强度控制在1 000 μmol·m-2·s-1,参比室内的CO2浓度分别设定在0、50、100、200、400、600、800、1 000、1 200、1 500、1 800、2 000 μmol·m-2·s-1,共有12 个CO2的浓度梯度,重复测定3 次。
1.3 数据处理与分析
数据整理采用Excel 2020 软件,数据分析采用SPSS 20.0 软件,多重比较采用Duncan 方法,显著性水平P≤0.05,极显著水平P≤0.01。利用光合计算软件对实测的光合数据进行拟合,得到各项目的光合计算参数,进而对7 个黄精种间的光合参数进行分析[15]。
2 结果与分析
2.1 7 种黄精属植物的净光合速率日变化
7 种黄精属植物均不存在午休现象,总体走势相近,7 种黄精属植物中点花黄精与黄精的净光合速率高于其余5 种,其净光合速率日变化呈明显的单峰曲线。上午随着外界光照辐射强度的增强黄精属植物的净光合速率迅速增加,下午随着外界光照辐射强度的减弱而随之降低。
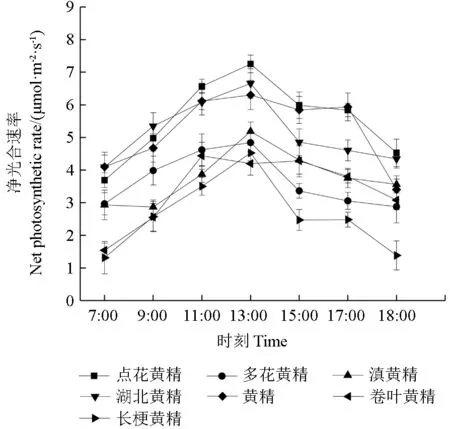
图1 7 种黄精属植物的净光合速率日变化Fig.1 Daily changes of net photosynthetic rate of seven Polygonatum plants
2.2 7 种黄精属植物的叶片叶绿素含量差异
由表1可以看出,7 个黄精属植物种间的叶片叶绿素含量差异极显著(F=23.3,P<0.01)。多重比较发现,黄精的叶片叶绿素含量最高,为1.83 mg·g-1,显著高于其它种,卷叶黄精的叶片叶绿素含量最低,为1.07 mg·g-1,显著低于其它种,其余黄精属植物叶片的叶绿素含量在1.25~1.54 mg·g-1之间。
2.3 7 种黄精属植物的水分利用率差异
表1所示7 个黄精属植物种间的水分利用效率差异极显著(F=11.33,P<0.01),其中点花黄精(5.13 μmol·mol-1)和长梗黄精(5.05 μmol·mol-1)的水分利用效率显著高于其它种,具有更好的抗旱性能,黄精(3.47 μmol·mol-1)、卷叶黄精(3.42 μmol·mol-1)和多花黄精(2.50 μmol·mol-1)亦显著高于湖北黄精(1.94 μmol·mol-1)和滇黄精(1.83 μmol·mol-1),其中滇黄精的水分利用率最低,其抗旱性较差。
2.4 7 种黄精属植物种间的气孔导度差异
表1显示7 种黄精属植物种间的气孔导度间差异不显著(F=0.925,P>0.05),在7 种黄精属植物中滇黄精的气孔导度最高,为0.122 mmol·m-2·s-1,湖北黄精的气孔导度最低,为0.737 mmol·m-2·s-1,所有种均分布于0.737~0.122 mmol·m-2·s-1之间,种间均不存在显著性差异。

表1 7 种黄精属植物光合指标参数的比较†Table 1 Comparison of photosynthetic index parameters of seven Polygonatum plants
2.5 7 种黄精属植物种间的胞间CO2 浓度差异
表1中7 种黄精属植物种间的胞间CO2浓度不存在显著差异(F=0.638,P>0.05),其中点花黄精的胞间CO2浓度最高(352.6 μmol·mol-1),长梗黄精的胞间CO2浓度最低,为312.6 μmol·mol-1,种间不存在显著差异。
2.6 7 种黄精属植物种间的蒸腾速率差异
表1所示为7 种黄精属植物种间的叶片蒸腾速率差异显著(F=21.0,P<0.01),其中湖北黄精和滇黄精的叶片蒸腾速率最高,分别为2.92和2.43 mol·m-2·s-1,显著高于其它种,两者水分利用率最低;长梗黄精的叶片蒸腾速率最低,为0.62 mol·m-2·s-1,显著低于其它种,其水分利用率最高,试验结果表明黄精的水分利用率与叶片蒸腾速率存在显著的负相关关系。
2.7 7 种黄精属植物的光响应曲线及其相关参数
从图2可以看出,7 种黄精属植物的叶片Pn值对光合有效辐射的响应曲线存在差异。7 种黄精属植物的Pn值在光照强度为0~500 μmol·m-2·s-1范围内会随着PAR 的增强,Pn也随之迅速增加。PAR 在500~1 000 μmol·m-2·s-1范围内,Pn上升趋势缓慢。在PAR 在1 000~2 000 μmol·m-2·s-1范围内,7 种黄精属植物的Pn的变化趋势相近,均呈下降趋势,点花黄精和卷叶黄精比较稳定,下降速度较慢,其余5 种黄精属植物保持持续下降趋势,下降速度较快,表明点花黄精和卷叶黄精对光照的利用区间较广。
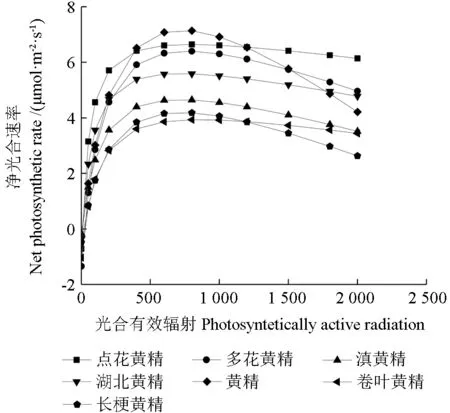
图2 7 种黄精属植物的光响应曲线Fig.2 Light response curves of seven species of Polygonatum
根据表2的多重比较发现多花黄精的暗呼吸速率最高,卷叶黄精的光补偿点最高。7 种黄精属植物最大净光合速率在4.03~7.15 μmol·m-2·s-1之间,黄精的最高为7.15 μmol·m-2·s-1,卷叶黄精的最低为7.99 μmol·m-2·s-1;光饱和点在709.44~861.85 μmol·m-2·s-1之间,其中卷叶黄精最高,表明其利用强光的能力最强,湖北黄精的最低。光补偿点在3.66~13.72 μmol·m-2·s-1之间,其中卷叶黄精的光补偿点最高为13.72 μmol·m-2·s-1,滇黄精的最低为3.66 μmol·m-2·s-1,表明滇黄精更适应弱光环境。
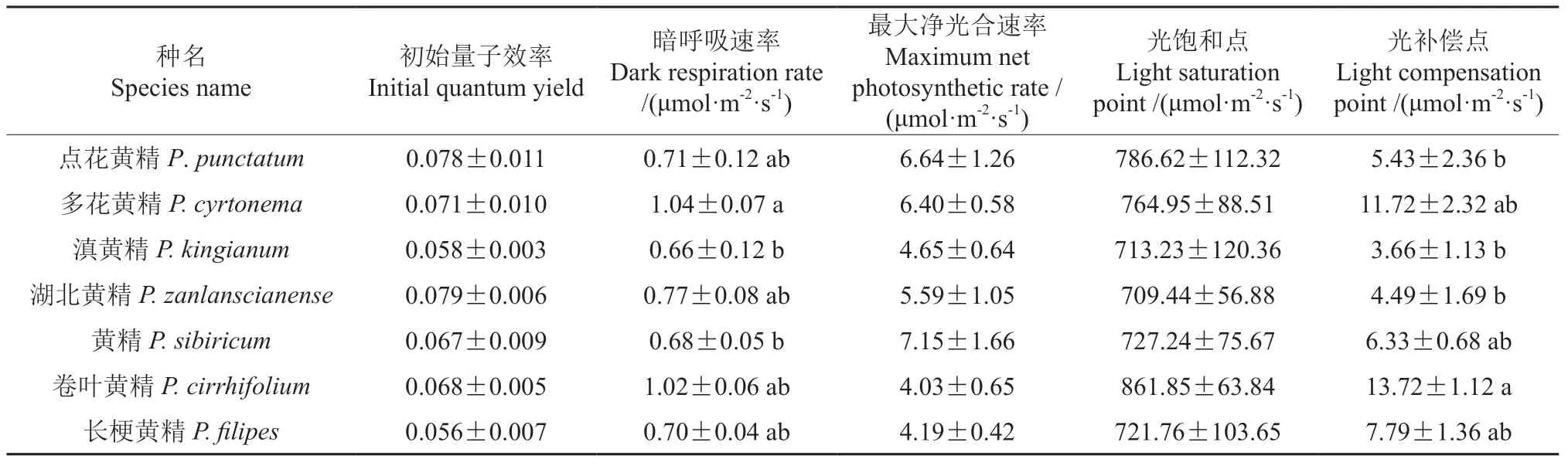
表2 光响应曲线相关参数Table 2 Related parameters of light response curve
2.8 7 种黄精属植物CO2 响应曲线
从图3可以看出,7 种黄精属植物的叶片Pn值对CO2的响应曲线趋势一致。CO2浓度在0~600 μmol·mol-1范围内,随着CO2浓度的增加,净光合速率迅速增加。CO2浓度在600~1 000 μmol·mol-1范围内,净光合速率增加速率变慢。CO2浓度在1 000~2 000 μmol·mol-1范围内,净光合速率缓慢增加,不同种间表现略有差异。
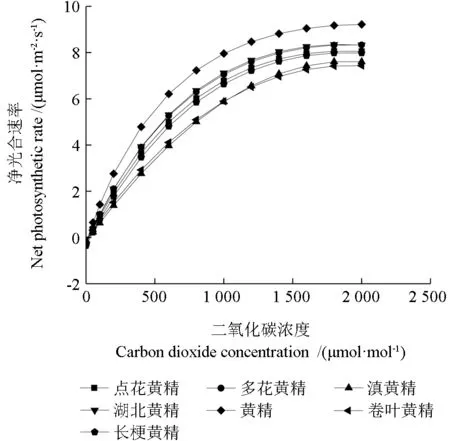
图3 7 种黄精属植物CO2 光响应曲线Fig.3 CO2 light response curves of seven species of Polygonatum
2.9 光合作用参数间的相关性
通过对7 种黄精属植物的光合参数的平均值间做相关性分析可以看出(表3),黄精属植物叶片净光合速率分别与叶片的胞间CO2浓度和水分利用率存在显著的正相关关系,相关系数分别为0.691 和0.905(P<0.05);叶片的气孔导度和蒸腾速率间也存在显著的正相关关系,相关系数为0.712(P<0.05);胞间CO2浓度与叶片蒸腾速率间存在显著正相关关系(R2=0.64,P<0.05)。
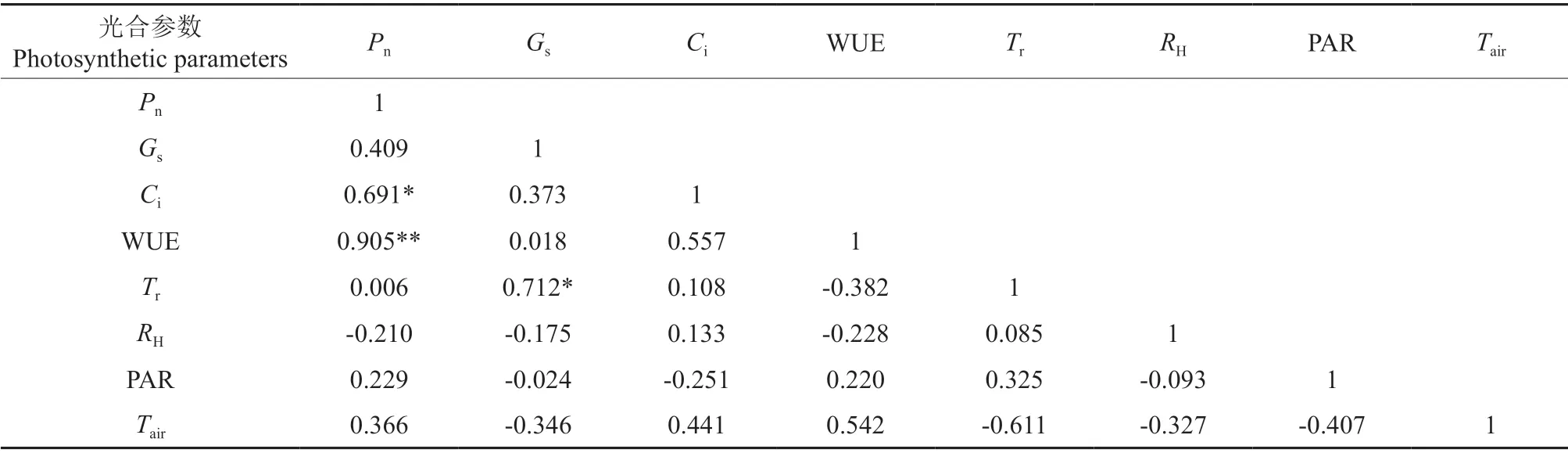
表3 光合作用参数间的相关性†Table 3 Correlation among photosynthesis parameters
2.10 7 种黄精属植物的光合特性评价
光合速率的大小受内外多种因素的影响,试验选取了7 种黄精属植物光合特性的5 个主要指标:净光合速率、气孔导度、蒸腾速率、胞间CO2浓度和水分利用效率的平均值,在SPSS 数据处理系统中,选择相似矩阵和最远邻元素聚类方法,聚类成3 类[16-17]。由聚类结果结合表1~2 中7种黄精属植物光合速率的大小可以得出,7 种黄精可分为3 类:多花黄精、黄精和滇黄精分为一类,其综合光合能力最好;湖北黄精、卷叶黄精和点花黄精同属一类,综合光合能力次之;长梗黄精是其中综合光合能力最差的一类。
3 结论与讨论
1)7 种黄精属植物的叶片净光合速率与胞间CO2浓度和叶片的水分利用率呈显著性正相关。7 种黄精属植物叶片叶绿素含量存在显著性差异,但其净光合速率与叶绿素含量之间并无显著相关关系,这一点在其它作物上也有相似的报道[18]。植物叶片的净光合速率是评价植物光合作用强弱的一个重要指标,影响叶片净光合速率的因素有两个方面,一方面是叶片本身,如叶龄、叶形、气孔阻力、蒸腾速率、叶绿素含量及叶向等均对叶片光合速率产生影响;另一方面主要受外界环境,如温度、光照强度、空气湿度等的影响[19-20]。对净光合速率和光合色素以及其它指标进行相关性分析,找出与净光合速率相关的指标,可以为生产中提高植物的光合能力提供一定的理论依据[21-22]。
2)7 种黄精属植物中,点花黄精和长梗黄精的水分利用效率相对较高,叶片蒸腾速率相对较低。水分利用效率是指单位面积水分消耗所能同化的光合产物量[23],是一个较为稳定的衡量碳固定和水分消耗比例的指标。研究表明,干旱胁迫下的小紫珠、日本荚蒾幼苗的光合速率都受到显著的影响[24-25]。通过测定WUE 能够阐明植物的气体交换特性以及抗旱性机理,从而选择出低耗水、生产效率高、抗性强的黄精属植物[26]。本试验条件下,7种黄精属植物的WUE 种间差异显著,表明不同种间水分利用能力差别较大,点花黄精和长梗黄精的水分利用效率较高,与其对应的叶片蒸腾速率较低,表明这两个种的耐旱性相对更强。
3)7 种黄精属植物的光饱和点接近阳生植物,光补偿点接近阴生植物,CO2饱和点高于正常外界浓度。7 种黄精属植物虽然喜阴,但是对于有效光合辐射有着比一般阴生植物更高的需求,种植中在提供阴凉环境的同时也应尽量满足其光照需求[27]。从7 种黄精属植物的CO2饱和点来看,均能适应较高浓度的CO2水平。所以,随着大气CO2浓度的增加,如果在其它条件满足的条件下,将更有利用黄精的生长和产量的提高。
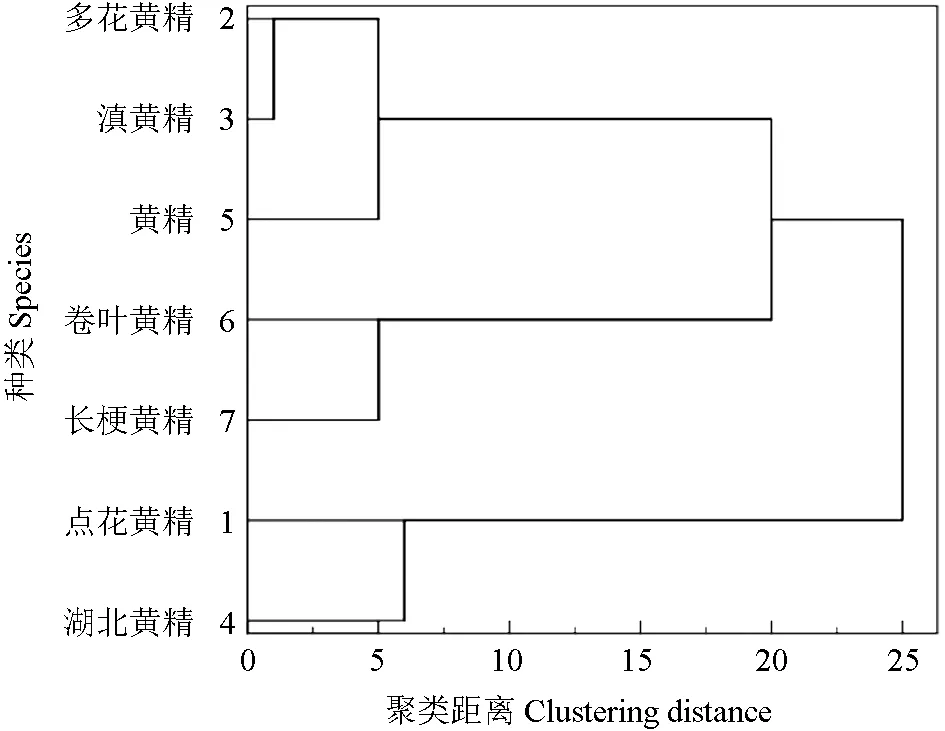
图4 光合特性聚类分析图Fig.4 Cluster analysis of photosynthetic characteristics
光合指标的聚类分析显示,7 种黄精属植物可聚成3 类,其中多花黄精、黄精和滇黄精归为光合性能最优类。
多花黄精、黄精和滇黄精是《中国药典》认定的药用黄精正品,其种植历史悠久,结合本研究结果,可以加大推广多花黄精、黄精与滇黄精的种植面积,并加强保护多花黄精、黄精与滇黄精的野生种质资源。点花黄精、湖北黄精和卷叶黄精的光合性能次之,应结合产量、品质等综合特性合理进行种植。长梗黄精的光合特性较差,应结合其它性状综合考虑是否适宜栽培。
猜你喜欢
西南农业学报(2021年7期)2021-08-23
今日农业(2021年9期)2021-07-28
节水灌溉(2021年5期)2021-05-26
中国农业科技导报(2020年12期)2020-03-15
小聪仔(幼儿版)(2020年11期)2020-02-01
新课程·下旬(2017年7期)2017-08-14
西藏大学学报(自然科学版)(2015年1期)2015-12-25
西藏科技(2015年9期)2015-09-26
西北林学院学报(2015年5期)2015-01-03
植物营养与肥料学报(2011年2期)2011-10-26
