
茉莉酸调控植物应对逆境胁迫作用的研究进展
2021-08-10 01:31毛佳昊熊晓辉卢一辰
生物加工过程 2021年4期
毛佳昊,熊晓辉,卢一辰
(南京工业大学 食品与轻工学院,江苏 南京 211800)
植物的生长发育离不开外界环境,承阳光雨露之恩泽,受风雷禽虫之折损。在自然环境下,凡是对植物生存或生长不利的环境因子统称为逆境或胁迫。在长期的进化过程中,植物对不利环境发展出了适应性和抵抗力,称之为耐逆性或抗逆性。植物遭受机械损伤、植食性动物的攻击或处于极端条件(干旱、高盐、极端天气、重金属、农药等)下时,一些植物激素作为信号分子大量累积,并诱导机体发生复杂的生物化学反应来激活自身免疫系统,以应对环境的生物或非生物胁迫。
茉莉酸(jasmonic acid,JA)作为其中一种重要的“胁迫激素”,是一种已被公认的新型植物生长调节剂。目前已有研究表明,茉莉酸类化合物与植物抗逆性密切相关,能调控与抗逆有关的基因表达,从而缓解植物受到生物或非生物胁迫[1]。至此,茉莉酸受到了逆境生物学家的广泛关注和研究。
本文中,笔者综述了茉莉酸的发现过程、生物功能、合成路径以及对病虫害、盐、干旱、重金属、农药胁迫等生物及非生物胁迫的作用。
1 茉莉酸简介
茉莉酸是一类脂肪酸的衍生物(图1),是包括游离态的茉莉酸及其衍生物和活性前体物质12-氧-植物二烯酸(12-oxo-phytodienoci acid,OPDA)等的总称。其衍生物主要有2种,分别为茉莉酸-氨基酸(如JA-Ile)和茉莉酸甲酯(MeJA)。Wasternack等[1]发现,茉莉酸及其衍生物在植物发育和对生物及非生物胁迫反应过程中发挥重要的作用。然而,与生长素、细胞分裂素、赤霉素、乙烯和脱落酸五大经典的传统类植物激素相比,茉莉酸及其衍生物是一类较新步入研究者视野的植物激素[2]。
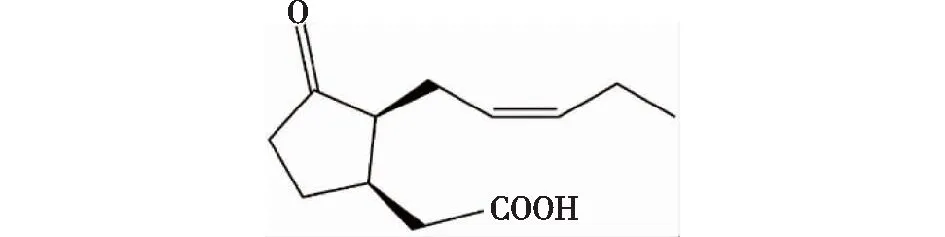
图1 茉莉酸的结构Fig.1 The structure of JA
1962年,Demole等[3]从素馨花的香精油中发现并分离得到了具有芳香味的物质茉莉酸甲酯。1971年,Aldridge等[4]从龙眼焦腐病菌培养基中分离出了茉莉酸,并发现龙眼焦腐病菌产生的这种游离态茉莉酸能够抑制植物生长。随后,科学家陆续从芸豆未成熟的种子中、日本板栗新鲜叶片、虫瘿和葫芦果种子中分离得到游离态的茉莉酸[5]。直到1980年茉莉酸类化合物的生物活性才慢慢被揭开。Ueda等[6]从苦艾中分离出(-)-methyl jasmonate并在艾草体内证明其可以促进衰老,进一步比较了(-)-methyl jasmonate和(+/-)-methyl jasmonate2种异构形式的茉莉酸甲酯生物活性,发现只有(-)-methyl jasmonate是具有促衰老的生物活性。次年,Dathe等[7]分离了(-)-methyl jasmonate异构体并发现其能抑制蚕豆种皮的发育,该研究结果首次报道了茉莉酸能抑制植物生长发育。同年,Yamane等[8]通过化学合成得到JA以及JA类似物,并发现其可以抑制水稻生长。茉莉酸类化合物的生物学功能不断得到挖掘,目前已发现其参与调控了植物生长发育的各个方面,如:花粉成熟,根生长,花青素累积,表皮毛的发生,植物的育性,气孔开闭、开花时间、根长抑制、块茎形成、种子萌发、果实成熟和植物的衰老等[9-15]。
2 植物体内茉莉酸的生物合成
经Wasternack等[1]研究发现,大部分植物体内合成茉莉酸的途径都比较相似,其合成可以分为2个阶段(图2)。第1个阶段发生在质体中,JA生物合成的脂肪酸底物是从叶绿体膜的半乳糖脂释放的α-亚麻酸(α-LeA)。α-LeA的氧合作用是JA生物合成的最初步骤,氧原子必须通过脂氧合酶(LOX)插入C13位置;之后在丙二烯氧化合酶(AOS)及丙二烯氧化物环化酶(AOC)的作用下生成12-氧-植物二烯酸(OPDA)。第2个阶段发生在过氧化物酶体中,OPDA在OPDA还原酶(OPR3)及3次β-氧化作用下,形成(+)-7-iso-JA,最终通过异构化作用,(+)-7-iso-JA很容易转化成(-)-JA,其合成过程如图2所示,其中AOS、AOC、OPR3为茉莉酸合成限速酶[1]。
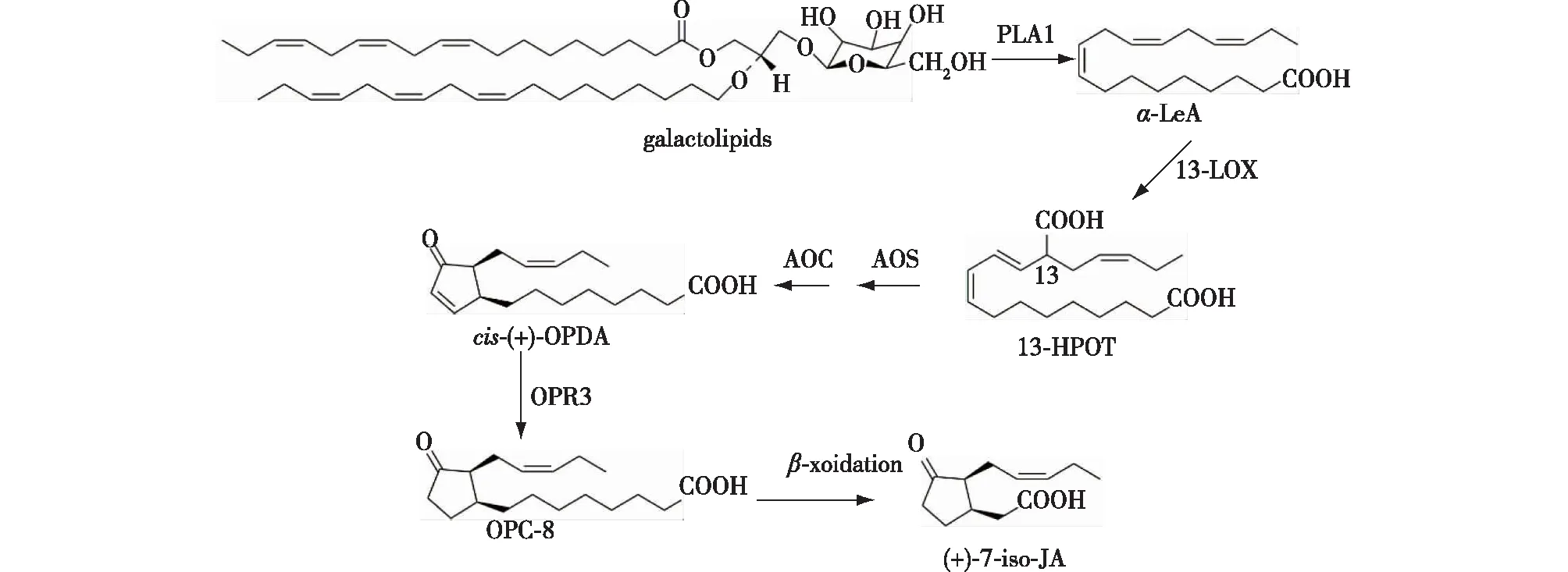
图2 茉莉酸合成途径[1]Fig.2 Jasmonic acid synthesis pathway[1]
3 茉莉酸的抗逆机制及信号传导
植物不断整合环境信号,以优化繁殖成功的方式调节其生长、发育和新陈代谢。除了可以预测诸如光照和温度之类的昼夜线索外,植物还能够感知和应对不可预测的环境影响,如缺水、感染、寄生等。这些选择性压力塑造了信号网络的出现,在这种网络中,小分子激素在将胁迫感知与复杂的转录反应相关的中心作用中发挥了重要作用,从而促进了植物在不利环境中的适应能力[16]。
许多研究表明,茉莉酸、茉莉酸甲酯和其他茉莉酸盐是阻止植物天敌专门的代谢物和蛋白质的有效激发因子。JA-Ile核心信号通路中的模块可以将各种环境影响与多种生理表现联系起来(图3)[17]。诱导信号包括微生物、草食动物和与损害相关的分子模式(分别为MAMP、HAMP和DAMP),它们来自生物攻击、受损的植物细胞和其他环境胁迫。这些信号被细胞表面的模式识别受体识别,从而触发JA-Ile的合成。JA-Ile通过E3泛素连接酶SCFCOI1和26S蛋白酶体的作用促进多种JAZ蛋白的降解(以JAZx-z为例)。JAZ的降解减轻了与JAZ相互作用的转录因子(以TFa-c为例)的抑制,该转录因子控制着涉及生长、发育以及对生物和非生物胁迫耐受性的各种生理输出响应。保守的核心途径可以通过进化来重新利用,以将其他输入信号与特定的转录反应联系起来[17]。
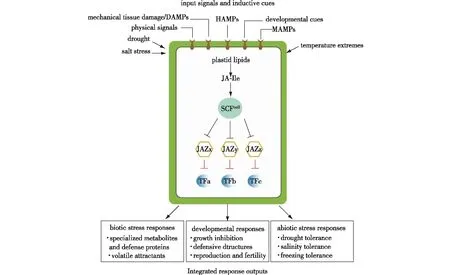
图3 核心JA-Ile信号传导与抗逆过程[17]Fig.3 The stress resistance and core JA-Ilesignal pathway[17]
4 茉莉酸对植物逆境胁迫的影响
茉莉酸还在植物对生物和非生物胁迫的抗性反应中起重要作用,其作为内源信号分子参与包括力学损伤、昆虫侵害、腐蚀性病原菌侵袭等生物胁迫和低温、干旱、盐胁迫以及重金属胁迫的抗性反应过程[1]。
4.1 生物胁迫
4.1.1 茉莉酸与植物的抗虫性
JA对植物抗虫性的研究主要集中在以下4个方面。
1)外源JA诱导植物产生行为干扰物质。Lawrence等[18]利用外源JA处理落叶松(Larixgmelinii),结果发现松针体内单宁含量显著增加。单宁可以通过与昆虫的唾液蛋白和消化酶结合,导致昆虫消化酶失活,从而干扰昆虫对植物蛋白的消化吸收;van Dam等[19]用JA处理番茄3 d后,番茄叶片中的尼古丁比对照组含量增加了2倍,取食其叶片的烟草天蛾幼虫发育减缓,死亡率显著升高。
2)外源JA诱导植物引诱寄生性天敌。植物在损伤或被植食动物攻击后会释放出大量的挥发性物质,而且所释放的挥发物无论是在种类还是在数量方面都发生了明显变化,这些变化会成为植食性动物的天敌实现寄主定位的指示物。有研究发现外源茉莉酸处理可诱导产生类似的挥发物,主要包括萜烯类、含有6个碳原子的醇、酮和酯类[20]。Meiners等[21]用1 μmol/L JA处理榆树(Ulmusminor),诱导产生的挥发性化合物吸引了榆叶甲(Xanthogalerucaluteola)的寄生性天敌啮小蜂(Oomyzusgallerucae),结果发现JA诱导处理的榆树上天敌数量显著比对照多。
3)外源JA诱导植物产生防御蛋白。茉莉酸类化合物诱导植物产生防御性蛋白,可刺激消化酶的过量分泌,经神经系统传递使昆虫产生厌食、拒食反应,导致昆虫发育不良;还调节植物蛋白酶抑制剂基因的表达,诱导蛋白酶抑制剂的合成。激酶、磷酸酶及调节蛋白酶等酶类在昆虫的生长发育过程中起着重要的调节作用,对植物喷施JA后会产生一些抑制这些酶活性的化学物质,进而抑制昆虫的生长发育,甚至导致植食性昆虫的死亡[22]。多酚氧化酶(PPO)是植物体内广泛存在的一种抗营养酶类,能够催化多酚化合物次生代谢,降低植物的营养价值,作为一种重要的防御性酶,能够被外源茉莉酸和茉莉酸甲酯诱导产生,阻止昆虫取食、抑制昆虫生长发育[23]。黑杨(Populusdeltoids)叶片经茉莉酸甲酯熏蒸后,酚类物质含量显著增加,体内PPO活性也显著升高,将MeJA处理过的叶片饲喂杨扇舟蛾(Closteraanachoreta)的幼虫,发现幼虫的生长发育明显被抑制[24]。Constabel等[25]应用25 μmol/L MeJA处理18种植物,诱导其PPO活性,结果表明,被诱导的烟草(Nicotianatabacum)和杨树(Populustrichocarpa)与未处理的相比,PPO活性分别上调了22.9倍和5.4倍。Thaler 等[26]应用1.5 mmol/L外源JA处理番茄,结果番茄PPO活性显著提高,减少了鳞翅目幼虫、甲虫、蚜虫和蓟马等4类昆虫的危害。Chen等[27]用JA喷施菊花幼苗不同部位,观察各部位对西花蓟马的抗性,结果表明在叶尖上局部施用JA提高了叶片PPO活性,且植物营养成分含量下降,取食叶片的西花蓟马的体质量明显低于对照组。
4)外源JA诱导行为干扰物质。Kessler等[28]利用外源MeJA诱导烟草对番茄天蛾(Manducaquinquemaculata)的抗性。结果表明,用200 μg/L MeJA诱导的每一株烟草挥发出的有机化合物,对番茄天蛾成虫产卵行为驱避作用极强,处理过的烟草上着卵量比未处理的低90%,但这种起驱避作用的化合物并未鉴定出。刘勇等[29]用5 mmol/L外源茉莉酸诱导小麦对麦长管蚜(Sitobionavenae)和禾谷缢管蚜(Rhopalosiphumpadi)的取食防御机制,应用电穿透(electrical penetration graph)技术测定发现,JA诱导后,2种蚜虫的非取食波次数和历时显著增加,说明JA诱导使小麦产生了防御反应,显著降低了蚜虫的取食程度。这与小麦被诱导产生维管液营养成分发生改变有关,驱避了蚜虫的取食。
4.1.2 茉莉酸与植物的抗病性
在诱导植物产生抗病性方面,向妙莲等[30]发现,MeJA可作为信号分子,参与植物对病原菌作出的应答并进行信息传递,激活植物体内活性状态的防御系统,诱导植物的抗性反应,进而提高植株抵抗病原菌侵害的能力。有研究利用JA合成途径的基因缺陷突变体aos、def1以及jar1证实了JA合成突变植株对番茄灰霉病菌、香蕉枯萎病菌和棉花黄萎病菌都表现出高易感性[31]。
Jia等[32]测定了葡萄果实发育过程中JA含量及其生物合成和信号相关基因的表达水平;进一步研究了MeJA在葡萄果实上的应用,表明MeJA能促进葡萄果实着色、软化和香气传播,特别是荚果、几丁质酶、β-1,3-葡聚糖酶和苯丙氨酸解氨酶等抗性物质的积累,并且在体外观察到MeJA抑制了番茄灰霉病菌的孢子萌发、胚芽管生长和菌丝延伸,提示MeJA保护果实免受霉菌感染。
4.2 非生物胁迫
4.2.1 茉莉酸与植物抗旱
干旱问题是影响作物生长并导致产量降低最为普遍的非生物胁迫因子之一。干旱胁迫影响植物的代谢和发育过程、损害膜系统、降低植物的光合作用,甚至造成植物死亡。植物本身对干旱胁迫有一定的承受能力,但对于超过一定范围的干旱胁迫,植物就失去了自身的调节能力。目前茉莉酸类化合物对植物的抗旱影响已有大量研究报道,其主要通过以下几个方面调节植物对干旱的胁迫:①调控植物气孔开闭和光合作用强度;②促使植物体内渗透调节物质含量升高;③调节植物抗氧化酶体系。
韩瑞宏等[33]对紫花苜蓿研究发现,缺水环境能够抑制植株的气体交换,使叶绿素含量降低,甚至破坏其结构。MeJA在调节气孔运动、维持叶绿素含量方面起到了关键作用。外源MeJA可减轻干旱胁迫下烤烟叶片叶绿素的分解[34];马超等[35]发现在干旱胁迫下喷施MeJA可有效促进β-胡萝卜素的积累,保护其他质体色素免受氧化降解,并通过调节气孔开闭度避免植株水分的散失,从而缓解了干旱胁迫对小麦叶片光合同化能力的不良影响,增强小麦的抗旱性。
脯氨酸和可溶性糖作为植物体影响巨大的渗透调节物质,在逆境条件下,可以通过降低细胞渗透势的方法来达到提高细胞吸水性的目的,使植物减轻不良环境带来的影响。李兆举等[36]研究发现,采用低浓度(0.25 μmol/L)外源MeJA浸种能够促进干旱胁迫下玉米种子的萌发以及幼苗正常生长,促进植物幼苗的脯氨酸含量升高,高浓度MeJA浸种则对种子萌发和幼苗生长产生明显的不良影响。在缺水状况下,植物会产生大量活性氧,细胞膜发生脂质过氧化,膜系统受到氧化伤害,导致植物组织损伤。Alam等[37]观察到干旱对油菜产生的氧化损伤,表现为脂氧合酶的活性和氧化应激、丙二醛和H2O2的水平升高,用0.5 mmol/L MeJA喷施油菜,提高了油菜体内的抗氧化酶体系活力,并且提高了作物的鲜质量和叶绿素含量。
4.2.2 茉莉酸与植物抗盐
盐害是目前制约农作物生产的主要逆境因素之一,土壤中较高的盐离子浓度可以改变土壤结构,降低土壤透气性和导水性,对植物造成渗透压胁迫,打破植物体内的离子和水势平衡,引起植物毒害,抑制植物生长发育,严重时甚至导致植物死亡。它对植物造成的损伤主要包括渗透胁迫带来的生理干旱和盐离子本身对植物的毒害。大量研究表明,JA通过影响气孔行为和调控蒸腾来调节植物对盐胁迫的反应。
Tani等[38]研究发现:盐胁迫诱导胁迫相关蛋白及茉莉酸合成基因的表达,水稻叶片与根中茉莉酸含量增加[38],耐盐品种茉莉酸含量高于盐敏感品种。Dombrowski等[39]研究发现,盐胁迫能够促进番茄中蛋白酶抑制剂的积累,而JA合成过程缺陷突变体def-1,在盐胁迫环境下,蛋白酶抑制剂的含量显著降低,表明JA参与盐胁迫下蛋白酶抑制剂积累的过程。MeJA预处理豌豆后也能够降低盐胁迫对CO2固定率、核酮糖-1,5-二磷酸羧化酶/加氧酶含量及活性、蛋白含量的抑制作用[40]。
近年来,对茉莉酸及衍生物诱导植物抗盐性的分子机制开展了不少探究。有研究发现,盐胁迫诱导多个物种的OPR1基因的表达,例如,水稻OsOPR1[41]、玉米OPR1和小麦TaAOC1基因[42]。在拟南芥中过表达玉米ZmOPR1基因发现,该过表达株系对渗透胁迫以及高盐胁迫的忍耐性相对于野生型而言有显著提高。Zhao等[43]在小麦中过表达TaAOC1基因后发现,植株内源性JA含量增加,耐盐性有显著提高。
4.2.3 茉莉酸与植物抗重金属
一些重金属元素,如,铁(Fe)元素是植物所必需的微量元素,对植物的生长起着很重要的作用。而大多数来自工业废料的重金属元素,如:镉(Cd)、汞(Hg)、铅(Pb)等,在机体内积累在一定阈值时,就会对植物的生长发育产生毒害作用。重金属主要通过以下几种途径对植物起到毒害作用:①抑制光合作用相关酶活性,降低光合效率;②取代植物必须营养元素和部分酶活性位点,导致酶丧失生理功能;③干扰其他离子的运输;④激发活性氧产生,造成植物细胞氧化损伤,继而影响植物生理代谢活动[44]。
有研究指出,外源茉莉酸甲酯可以诱导植物细胞防卫基因的表达,缓解重金属对植物的损害。陈俊[45]以秋燕幼苗为研究对象,对Cd胁迫下的秋茄幼苗使用MeJA并分时间段采样,测量叶绿素含量、丙二醛和抗氧化物质含量等指标,结果显示低浓度的MeJA可以有效缓解重金属镉对秋茄生长的抑制。Chen等[46]研究发现施加10 μmol/L MeJA和100 μmol/L JA,可以有效降低植物组织内金属硫蛋白(MT2)的表达,减少重金属向地上部位的运输,从而有效地缓解重金属的毒害作用,但当激素浓度过高时,本身也能够对植物产生伤害。
4.2.4 茉莉酸与植物抗农药
农药是一类用于防治农作物及农副产品的病虫草害及其他有害生物的化学试剂的总称,其中有些还广泛用于卫生、畜牧、水产、森林等方面病虫害的防治。虽然农药的使用防止了多种病虫害的发生,使农作物的产量有了极大的提高,但是目前过量、不规范的使用农药使人类健康面临严峻的挑战。
农药还通过产生有害的活性氧(ROS)对植物产生负面影响,从而导致植物细胞中的氧化胁迫[47],最终导致植物生长抑制,同时光合作用效率下降[48]。为了最大程度减少农药造成的氧化损伤,植物内部具有的防御系统(酶和非酶抗氧化防御系统)可在胁迫条件下被激活[49]。植物还可以通过酶介导的解毒系统对异种生物进行解毒,该系统包括诸如细胞色素P450酶、过氧化物酶、单加氧酶、羧酸酯酶和谷胱甘肽-S-转移酶等[50]。
茉莉酸可以通过调节植物的抗氧化防御系统,从而减少非生物胁迫条件下的氧化应激。Sharma等[51]用10 μmol/L的JA浸泡油菜种子,在受到吡虫啉(IMI)胁迫后,油菜幼苗酶促和非酶促抗氧化能力显著提高,并且JA的浸泡使油菜幼苗的IMI积累量显著降低。张书浩[52]以小麦为研究对象,研究异丙隆对小麦的毒性作用,表明外源添加茉莉酸甲酯可以有效缓解异丙隆对小麦的毒害作用,提高异丙隆胁迫下叶绿素的含量,缓解异丙隆对光合反应的抑制作用,同时茉莉酸甲酯降低了异丙隆对小麦的氧化胁迫以及在小麦体内的积累,并且外源添加MeJA可以促进土壤中农药异丙隆的降解。Kaya等[53]用45 μmol/L MeJA处理除草剂甲咪唑烟酸胁迫的烟叶,与对照组相比,施用JA的烟草叶片色素含量、抗氧化活性和植物激素水平增加,而丙二醛含量和除草剂残留量减少。
5 总结与展望
茉莉酸作为高等植物体内的一种内源生长调节物质,参与调控了植物大多数的生理生长过程,在植物的生长发育过程中起到了积极的作用。此外,茉莉酸又作为重要的逆境信号物质在调控多种植物逆境胁迫中起着重要作用。遗憾的是,学界目前对于茉莉酸如何调控某些具体的抗胁迫分子机制的了解不够详细。目前茉莉酸的成本较高,其对植物的抗逆研究多见于实验室,尚未有应用于大田的报道,了解茉莉酸的抗逆机制或可为绿色农业的发展提供依据。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
新世纪智能(数学备考)(2021年10期)2021-12-21
海洋通报(2021年5期)2021-12-21
农业资源与环境学报(2021年5期)2021-10-06
新世纪智能(数学备考)(2020年10期)2021-01-04
文苑(2020年12期)2020-04-13
意林·全彩Color(2019年11期)2019-12-30
意林彩版(2019年11期)2019-11-22
现代园艺(2018年3期)2018-02-10
中成药(2017年12期)2018-01-19
