
枯草芽孢杆菌胞外囊泡的分离及其对巨噬细胞免疫功能的影响
2021-09-14 07:55韩永先廖搏浪胡如久杨明明西北农林科技大学动物科技学院陕西杨陵712100
家畜生态学报 2021年8期
韩永先,林 华,廖搏浪,胡如久,苏 源,杨明明(西北农林科技大学 动物科技学院,陕西 杨陵 712100)
益生菌是一类能够促进肠道健康的活微生物[1]。许多研究表明,益生菌不仅可以预防肠道病原体定植,增强免疫系统[2-3],而且能够通过改善动物肠道形态结构和肠道微生态平衡来促进动物健康[4-5]。因此,全面发掘益生菌的功能对动物肠道健康有重要意义。枯草芽孢杆菌(Bacillussubtilis)是一种革兰氏阳性细菌,因其遗传背景清晰,在饲料加工中稳定性好,活性高,已成为目前应用最广泛的饲用益生菌制剂之一[6]。芽孢杆菌能够提高小鼠分泌IgG的含量,增强体液免疫功能[7],还能提高肠系膜淋巴组织分泌促炎性细胞因子和抗炎性细胞因子[8-9],从而平衡炎症反应,促进动物肠道健康。然而,关于B.subtilis如何引发免疫调节作用的详细机制尚未得到广泛研究。近年来,一些研究报道了细菌、真菌和寄生虫产生的细胞外囊泡(Extracellular Vesicles, EVs),具有介导细菌-宿主间的相互交流、呈递细菌抗原和调节宿主免疫功能等作用,因而受到广泛关注[10-11]。对细菌胞外囊泡的相关研究主要集中在革兰氏阴性菌分泌的外膜囊泡(OMVs)。在过去10年中,OMVs被认为是向靶细胞提供毒力和免疫调节因子的信号载体[12-13]。然而革兰氏阳性菌与革兰氏阴性细菌不同,其细胞壁主要由肽聚糖组成,没有外膜。但Dorward等[14]为革兰氏阳性菌释放胞外囊泡提供了首个证据。随后,人们陆续发现溃疡杆菌、金黄色葡萄球菌、李斯特氏菌和双歧杆菌等革兰氏阳性菌均可释放胞外囊泡[15-18]。近年研究发现,B.subtilis也能够释放EVs,经过蛋白组学分析,此菌株囊泡富含30种蛋白质,这些蛋白多数与囊泡的免疫调节相关[19]。巨噬细胞是源自骨髓前体的单核吞噬细胞,是先天性和适应性免疫反应的关键参与者[20],也是体外研究最常用的细胞模型之一。本研究通过检测不同浓度B.subtilis来源的胞外囊泡(BS_EVs)刺激巨噬细胞后产生的免疫细胞因子水平和免疫相关酶活水平,探究其对巨噬细胞免疫应答的影响,为B.subtilis的免疫调节机制提供理论依据。
1 材料与方法
1.1 细菌菌株与细胞株
枯草芽孢杆菌B.subtilis168为本实验室保存,RAW 264.7细胞购自中国科学院上海生命科学研究院细胞资源中心。
1.2 主要仪器设备
恒温摇床;高速冷冻离心机(美国Thermo);超速离心机(美国 贝克曼);Nanosight NS 300纳米颗粒跟踪分析仪(英国 马尔文);高速转盘式激光共聚焦(英国安道尔公司);场发射扫描电子显微镜(日本日立S-4800)。
1.3 细菌培养
B.subtilis168接种于LB固体培养基中,37 ℃过夜培养12 h,然后挑单克隆于LB液体培养基中在摇床进行37 ℃过夜培养。
1.4 细胞培养
RAW 264.7细胞系用60 mm培养皿在完全PRMI-1640培养基(含有10%胎牛血清,100 U/mL青霉素和100 μg/mL链霉素)中培养,培养条件为37 ℃和5%CO2,培养基每24 h更换1次,细胞每48 h传代1次。连续传代培养2次后,将1×105个细胞/mL的浓度接种到6孔培养板中,每孔加2 mL培养基(细胞悬液和新鲜完全培养基)继续培养 ,为后续试验做准备。
1.5 BS_EVs的分离与鉴定
将过夜培养好的B.subtilis168离心(12 000 g,20 min,4 ℃)除去菌泥,使用真空过滤装置将上清液通过0.45 μm孔径的过滤器过滤,可去除大颗粒,比如残留细菌和细胞碎片。然后通过装有100 kDa膜的Amicon超滤系统浓缩上清液。将浓缩好的上清液再次通过0.22 μm过滤器过滤以除去残留的细菌,最终利用超速离心(180 000 g,2 h,4 ℃)来制备EVs。为了使得到的EVs更加纯净、均一,采用OptiPrep(60% Density Gradient Medium,Sigma)密度梯度离心。密度梯度离心液浓度范围为10%~55%,即分为10层,每层溶液浓度相差5%。首先将EVs-PBS混合液与60%密度梯度液混合成55%的溶液,然后加入离心管底部,其余密度梯度液自下而上依次铺入离心管中,此时离心管中呈现10层界限较分明的溶液。操作完成后进行超速离心(200 000 g,16 h,4 ℃),离心完成后,将每一层溶液取出测其粒径大小及含量分布,取出试验目标物再离心1次,此时得到的沉淀就是纯化后的EVs。将EVs重悬于PBS中并储存在-80℃下用于随后的试验。通过纳米颗粒跟踪分析仪测定EVs的粒径分布,并且用扫描电子显微镜观察EVs的形态特征。利用BCA试剂盒(目录号T9300A,TaKaRa)测定EVs的蛋白质浓度(μg/mL)。
1.6 BS_EVs刺激RAW 264.7细胞试验
将6孔板中培养至单层的RAW 264.7细胞用PBS溶液洗涤3次并加入2 mL新鲜的完全培养基,试验组分别加入1、2、4、8和16 μg/mL的BS_EVs(即EVs1、EVs2、EVs4、EVs8、EVs16组),对照组加入等体积的PBS(PBS组)。置于37 ℃ 5%的细胞培养箱12 h。结束后用台盼蓝排除检测法测定细胞存活率,收集上清液并测定IL-10、TGF-β、TNF-α、IL-6和乳酸脱氢酶(LDH)活性,用10%的TritonX-100裂解细胞,测定酸性磷酸酶(ACP)、乳酸脱氢酶(LDH)、诱导型一氧化氮合成酶(iNOS)和超氧化物歧化酶(SOD)活性。细胞因子用ELISA试剂盒(武汉云克隆公司)检测,相关酶活均采用测试盒(购于南京建成生物工程研究所)检测。本试验设置6个处理,每个处理3个重复,共3次独立重复试验。
1.7 数据处理
试验数据采用Excel软件整理,one-way ANOVA进行方差分析,用Duncan法进行多重比较。数据均用“平均值±标准差”表示,P<0.05为差异显著。
2 结果与分析
2.1 BS_EVs的分离、纯化与鉴定
将超速离心后获得的BS_EVs进行密度梯度离心,采用纳米颗粒跟踪分析仪测定各密度层BS_EVs的浓度,计算各层BS_EVs相对丰度,结果(图1 A)显示BS_EVs主要集中在F3~F5,所以选择浓度梯度在20%~30%的密度层进行超速离心富集得到纯化的BS_EVs。纯化后的BS_EVs大小主要为85 nm,且其粒径范围在50~200 nm(图1 B)。在扫描电镜下(图1 C)可以看到,BS_EVs的形态大小不一,呈圆形或椭圆形,直径平均为50~100 nm,符合细菌囊泡的理论大小值。
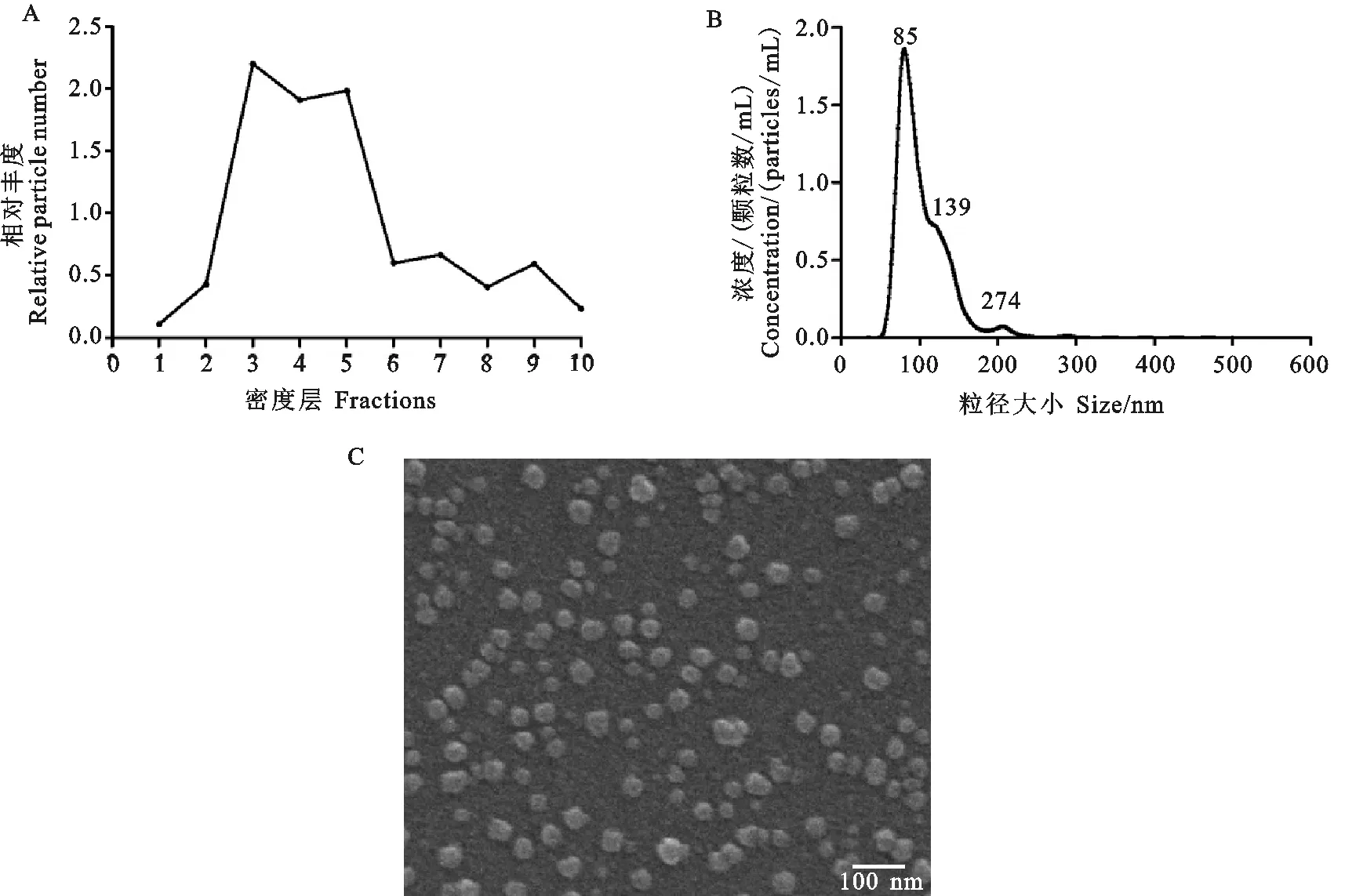
图1 BS_EVs的分离、纯化与鉴定A.密度梯度离心测定各层颗粒的相对丰度;B.纳米颗粒跟踪分析仪检测纯化的BS_EVs粒径分布; C.扫描电镜检测BS_EV形态。标尺为100 nmFig. 1 Isolation, purification and identification of BS_EVsA. The relative number of particles in each fraction determined by density gradient centrifugation; B. Size distribution of BS_EVs as determined by nanoparticle tracking analysis; C. BS_EVs were visualized using scanning electron microscope. Bar=100 nm
采用BCA法对纯化的BS_EVs进行蛋白定量,结果显示1×1010particles/mL的BS_EVs的蛋白含量为5.1 μg/mL。
2.2 BS_EVs对RAW 264.7细胞活性的影响
LDH是一种胞内酶,当细胞膜受损后就会被大量释放到胞外。通过采用台盼蓝染色试验和胞外LDH活性测定来检测BS_EVs对RAW 264.7细胞存活率的影响。由表1可知,与对照组相比,EVs1、2和4组细胞存活率无显著性差异(P>0.05),EVs8和16组细胞存活率显著降低(P<0.05)。EVs1、2和4组LDH活性无显著差异(P>0.05),而EVs8和16两组胞外LDH活性显著升高(P<0.05)。这表明较低剂量(本试验中的1、2和4 μg/mL)BS_EVs对RAW264.7没有毒性,而较高剂量(本试验中的8和16 μg/mL)可造成RAW 264.7细胞膜严重损伤,显著降低细胞活力。
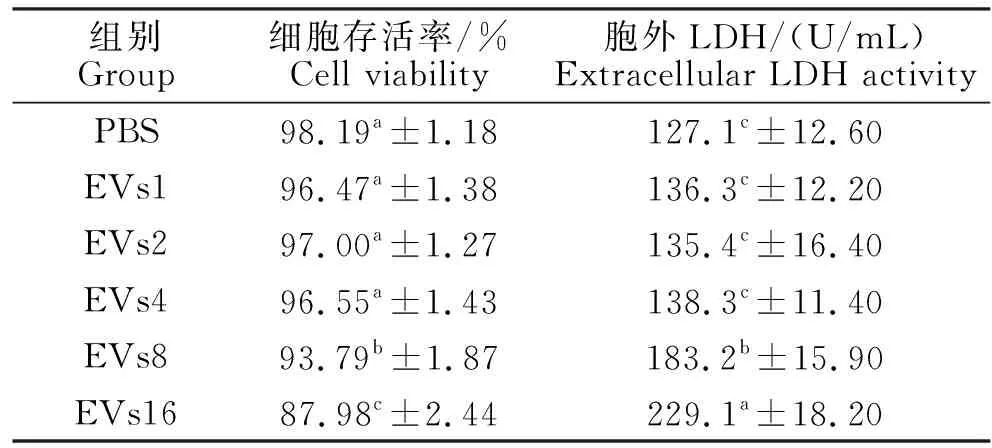
表1 BS_EVs对RAW 264.7细胞存活率和胞外 LDH活性的影响Table 1 Effects of BS_EVs on cell viability and extracellular LDH activity of RAW 264.7 cells
2.3 BS_EVs对RAW 264.7细胞免疫相关酶活性的影响
为了探究BS_EVs是否能够激活RAW 264.7细胞,检测了BS_EVs刺激后RAW 264.7细胞胞内酶活性的变化。由表2可知,与对照组PBS相比,5个BS_EVs组巨噬细胞内ACP活性显著增加(P<0.05),且EVs2和4组ACP活性最高;EVs1、2和4组胞内LDH活性显著高于对照组(P<0.05),EVs8组的胞内LDH活性无显著变化(P>0.05),而EVs16组的胞内LDH活性显著降低(P<0.05);各BS_EVs刺激组胞内iNOS活性显著高于对照组(P<0.05),且EVs2、4和8组iNOS活性显著高于其他两个组(P<0.05);与对照组相比,EVs1、2、4和8组胞内SOD活性无显著差异(P>0.05),而EVs 16组SOD活性显著降低(P<0.05)。
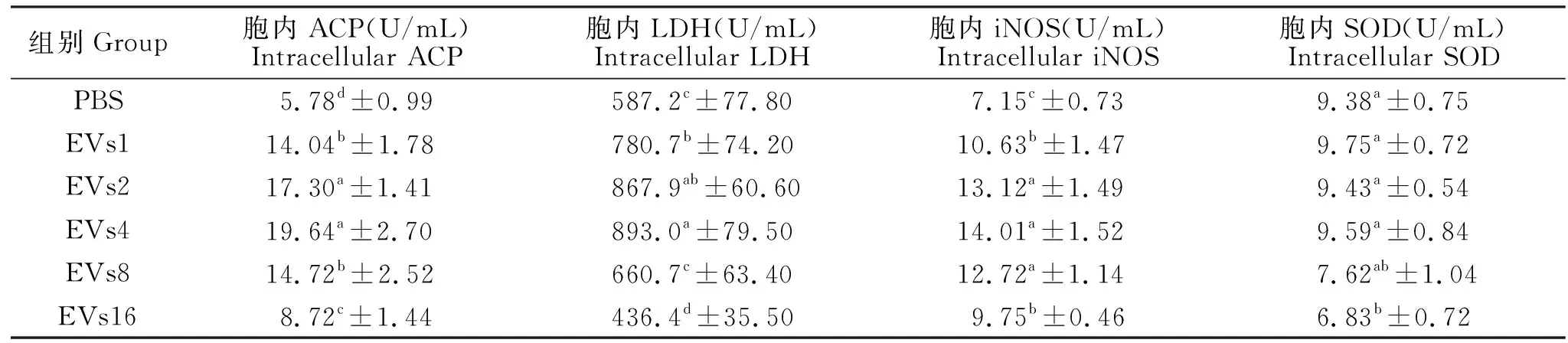
表2 BS_EVs对RAW 264.7细胞免疫相关酶活的影响Table 2 Effects of BS_EVs on ACP, LDH, iNOS and SOD activities in RAW 264.7 cells
2.4 BS_EVs对RAW 264.7细胞分泌细胞因子的影响
通过检测BS_EVs刺激后促炎性细胞因子(IL-6、TNF-α)和抗炎性细胞因子(IL-10、TGF-β)来评定巨噬细胞对BS_EVs的应答反应。由表3可知,BS_EVs刺激显著提高了促炎性细胞因子IL-6和TNF-α的分泌量(P<0.05),且随着BS_EVs剂量的增加而显著提高。BS_EVs刺激同时也显著提高了抗炎性细胞因子IL-10和TGF-β含量(P<0.05),其中EVs4、8组IL-10分泌量和EVs4组的TGF-β分泌量要显著高于其它BS_EVs刺激组(P<0.05)。这些数据表明,BS_EVs刺激同时激活了RAW 264.7细胞的促炎和抗炎应答反应。
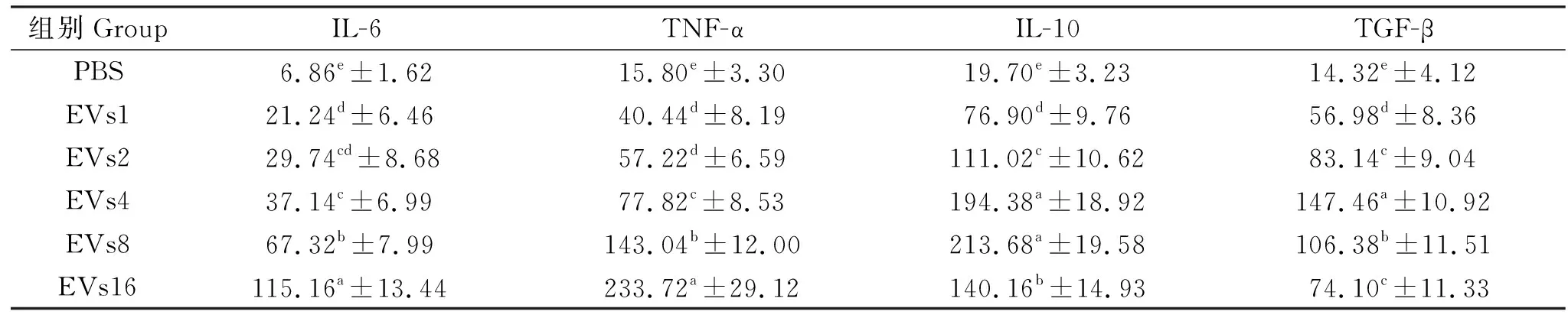
表3 BS_EVs对RAW 264.7细胞分泌细胞因子的影响Table 3 Effects of BS_EVs on the secretion of cytokines in RAW 264.7 cells pg/mL
3 讨 论
3.1 BS_EVs的分离、纯化和鉴定
大多数细胞,包括真核生物、革兰氏阴性和革兰氏阳性菌以及古生菌,都能分泌纳米大小的囊泡[21-22]。本试验研究的是革兰氏阳性菌枯草芽孢杆菌168来源的胞外囊泡,电镜观察发现这类囊泡的粒径大小在50~100 nm,此结果与前人的报道结果类似[19]。而囊泡分离方式的差异可能造成其大小和形态的不同。胞外囊泡的分离方式多样,包括超速离心法、超滤法(基于粒径大小)、膜亲和试剂盒法、沉淀法和微流术法[23],且都是针对真核细胞,对于原核细胞囊泡的分离没有系统的报道。本研究经过长时间的方法摸索,用超速离心法分离胞外囊泡,主要操作步骤参考Prados-Rosales等[24]的研究以及结合B.subtilis168本身生长特点总结出一套较完整的分离方法。其中,BS_EVs的纯化采用密度梯度离心法,此方法可以将不同粒径大小的胞外囊泡归类,将不同密度层的EVs相对丰度计算出来。本试验结果显示BS_EVs主要集中在20%~30%密度层,这与真核细胞分泌外泌体情况相似[25]。密度层相近的EVs的粒径大小和形态也较相似,因此从扫描电镜的结果可以看到,枯草芽孢杆菌菌株168分泌的囊泡浓度较大,且粒径大小比较均一。然而,由于细菌囊泡生物特性复杂导致其纯化方法耗时费力,如何快速高效的得到纯净的胞外囊泡还需后期继续研究。
3.2 BS_EVs对RAW 264.7细胞活性的影响
由于安全是益生菌的基本要求,所以安全应该是评估益生菌时的第一个项目。该研究使用台盼蓝排斥试验和乳酸脱氢酶毒性试验来研究BS_EVs对巨噬细胞活力的影响。台盼蓝试验结果表明低浓度EVs对巨噬细胞没有明显毒性作用,对细胞的生长来说是安全的。而高浓度EVs有降低细胞存活率的现象,此结果与EVs对巨噬细胞胞外LDH活性的影响一致。细胞膜通透性的增加总是伴随着细胞死亡[26],这将导致LDH的渗漏[27]。LDH是一种稳定的胞质酶,存在于所有细胞中[28],如果细胞膜受损,LDH将释放到胞外空间。因此,胞外LDH含量的增加是测量细胞膜损伤的指标之一[29]。本试验结果表明EVs8和16严重破坏巨噬细胞的细胞膜完整性,导致LDH大量流向胞外,而低浓度BS_EVs较好的维持了巨噬细胞的正常代谢状况。
3.3 BS_EVs对RAW 264.7细胞免疫相关酶活的影响
巨噬细胞(MΦ)是初始免疫系统中的主要参与者,从胃肠道和呼吸道黏膜进入机体的病原微生物首先遇到的免疫细胞是粘膜下组织的MΦ。MΦ组成性表达多种PRR,病原微生物通过与PRR作用而粘附在MΦ表面形成吞噬体,然后与溶酶体融合形成溶酶体酶,随即在多种溶酶体酶的作用下,病原体微生物被杀死[30]。所以,溶酶体酶是MΦ吞噬和消化异物的重要组成系统。ACP是一种水解酶,存在于溶酶体中,是其标志酶,所以ACP的升高反映了MΦ被激活的程度[31]。LDH是一种糖酵解酶,存在于MΦ胞质中,其功能是催化乳酸脱氧成为丙酮酸或丙酮酸还原为乳酸,在MΦ吞噬过程中其能量来源主要通过糖酵解途径,糖酵解过程中产生的乳酸使细胞胞内pH下降有利于溶酶体酸发挥作用[32],故其活性通常也被认为是被激活的标志之一[33-34]。本研究结果发现各组EVs显著升高巨噬细胞胞内ACP活性,低浓度组EVs组显著提高胞内LDH活性,表明低浓度组EVs均能激活MΦ的吞噬功能;而EVs 16组显著降低胞内LDH活性,表明高浓度EVs可能破坏了细胞膜完整性,导致LDH外泄。免疫系统中的巨噬细胞可被活化而引起表达,进而产生高量的NO,是巨噬细胞阻杀微生物及癌细胞的主要武器。Han等[35]研究发现,敲除iNOS基因的小鼠对多种胞内病原菌高度敏感。本试验结果表明各EVs组均能提高巨噬细胞iNOS活性,且EVs1、2和4组效果最佳,说明BS_EVs能活化巨噬细胞诱导表达iNOS,从而提高巨噬细胞抗菌能力。另外,EVs16组SOD活性显著降低,其他各组无显著差异,说明BS_EVs对巨噬细胞的抗氧化酶的产生无影响,且高浓度EVs组可能会减弱巨噬细胞的抗氧化应激。
3.4 BS_EVs对RAW 264.7细胞分泌细胞因子的影响
大多数炎症反应是先天免疫的结果,是通过巨噬细胞,多形核白细胞和肥大细胞激活促炎信号级联引发的[36]。细胞因子是由细胞产生的小分子可溶性蛋白质,它们主要调节宿主的免疫反应,造血功能和炎症反应,其中参与炎症反应的细胞因子分为2种主要类型:促炎因子(TNF-α、IFN-γ 、IL-1、IL-2、IL-6、IL-8、IL-12)和抗炎因子(IL-4、IL-10、IL-13)[37]。在本试验中,未加BS_EVs刺激时,巨噬细胞分泌的细胞因子含量很低,试验处理组用EVs刺激后,巨噬细胞分泌促炎性细胞因子IL-6和TNF-α的水平随着EVs浓度的升高而升高。同时,各浓度EVs也显著促进了抗炎因子IL-10和TGF-β的产生,IL-10是II型细胞因子,被认为是介导免疫抑制的细胞因子[38]。TGF-β是Treg相关细胞因子(负调控)介导免疫抑制或免疫无能[39]。此现象表明,BS_EVs同时增加了促炎因子和抗炎因子的产生,平衡了炎症反应,维持了免疫稳态。这与有些研究报道结果一致[11]。另外,EVs16组刺激细胞产生促炎因子的含量高于EVs4和8组,但是抗炎因子含量却低于EVs4和8组,说明高浓度BS_EVs极大增强了巨噬细胞的促炎反应。
4 结 论
低剂量(1、2和4 μg/mL)BS_EVs对RAW 264.7细胞活力没有显著影响,而较高剂量(8和16 μg/mL)显著降低细胞活力。适量BS_EVs(2和4 μg/mL)可提高RAW 264.7细胞ACP、LDH活性和NO信号通路,即提高了免疫反应相关酶活。
BS_EVs可同时增强RAW 264.7细胞的促炎反应和抗炎反应,本试验中低剂量组BS_EVs对抗炎反应的增强程度大于促炎反应,更有利于增强巨噬细胞的抗炎功能,高剂量组则相反。因此,适量BS_EVs可通过提高巨噬细胞免疫相关酶活、调节细胞因子分泌量来增强其免疫功能。
猜你喜欢
南京航空航天大学学报(2022年4期)2022-08-30
昆明医科大学学报(2021年12期)2021-12-30
中国食用菌(2021年10期)2021-11-04
现代临床医学(2021年4期)2021-07-31
昆钢科技(2021年6期)2021-03-09
昆明医科大学学报(2021年1期)2021-02-07
汽车零部件(2020年9期)2020-09-27
中国民族民间医药·上半月(2016年12期)2017-01-11
考试周刊(2016年90期)2016-12-01
浙江中医杂志(2004年5期)2004-03-09
