
甘露寡糖对养殖鱼类生长性能和非特异性免疫的研究进展
2021-12-18 02:28翟浩杰任同军王福强韩雨哲
安徽农业科学 2021年23期
翟浩杰 任同军 王福强 韩雨哲
摘要 由于抗生素的使用被限制,新型绿色饲料添加剂成为水产动物营养与饲料的研究热点,甘露寡糖作为一种功能性寡糖被认为是抗生素的替代品,研究发现,甘露寡糖能够促进动物生长,改善肠道健康并增强其非特异免疫功能。为此,综述了甘露寡糖对鱼类生长性能、肠道健康和非特异性免疫能力的影响,对其在水产养殖中的应用做出展望。
关键词 甘露寡糖;鱼类;生长性能;肠道健康;非特异性免疫
中图分类号 S 963.73 文献标识码 A
文章编号 0517-6611(2021)23-0017-05
doi:10.3969/j.issn.0517-6611.2021.23.005
Research Progress of Mannan Oligosaccharides on Growth Performance and Nonspecific Immunity of Cultured Fish
ZHAI Hao-jie1,REN Tong-jun1,2,WANG Fu-qiang1,2 et al
(1.College of Fisheries and Life Sciences,Dalian Ocean University,Dalian,Liaoning 116023;2.Key Laboratory of Applied Biology and Aquaculture of Northern Fishes in Liaoning Province,Dalian Ocean University,Dalian,Liaoning 116023)
Abstract Due to the limited use of antibiotics,new green feed additives have become the research hotspot of aquatic animal nutrition and feed.As a functional oligosaccharide,mannan oligosaccharide is considered as the substitute of antibiotics.Studies have found that mannan oligosaccharide can promote animal growth,improve intestinal health and enhance its non-specific immune function.Therefore,the effects of mannan oligosaccharides on growth performance,intestinal health and non-specific immunity of fish were reviewed,and its application in aquaculture was expected.
Key words Mannan oligosaccharides;Fish;Growth performance;Intestinal health;Non-specific immunity
作者简介 翟浩杰(1997—),男,陕西宝鸡人,硕士研究生,研究方向:水产动物营养与饲料。通信作者,副教授,博士,从事水产动物健康养殖和水产动物营养饲料学研究。
收稿日期 2021-03-25
近年来我国鱼类养殖业快速发展,高密度集约化的水产养殖导致的鱼类病害成為了影响渔业经济的重要因素之一,传统方法采用抗生素对病害进行防治,但由于抗生素等化学药物会对自然环境造成污染,而且容易在水产品中残留,严重影响食物安全和人类健康,因此,目前主要通过一些无毒无害无残留、结构稳定的饲料添加剂(寡糖、多糖、益生菌、氨基酸、维生素等)来提高动物的抗病能力和生产性能。
寡糖作为一种能源物质,自20世纪80年代被开发为饲料添加剂以来,一直是动物营养免疫方面研究的热点。寡糖又称低聚糖或寡聚糖,是由2~10个单糖单位经脱水缩合由糖苷键连接而成的小聚合体,寡糖又分为普通寡糖和功能性寡糖。甘露寡糖(mannan oligosaccharides,MOS)又称甘露低聚糖或葡甘露寡聚糖,是功能性寡糖的一类,是从酵母培养细胞壁中提取的一类新型抗原活性物质,广泛存在于魔芋粉、瓜儿豆胶、田菁胶及多种微生物细胞壁内。甘露寡糖是由几个甘露糖分子或甘露糖与葡萄糖通过α-1,2、α-1,3和α-1,6糖苷键组成的寡聚糖,易溶于水和极性溶剂,一般在生理pH和饲料加工制粒时较为稳定,能够保持其结构和功能的完整性。甘露寡糖作为一种新型绿色饲料添加剂,已经广泛应用于养殖动物饲料的开发,而作为饲料添加剂的甘露寡糖一般为二糖、三糖和四糖的混合物[1],研究表明甘露寡糖可以提高养殖动物的生产性能、改善肠道微生物区系以及提高免疫功能等[2-4]。大量研究表明,甘露寡糖具有促进鱼类生长性能,改善鱼类肠道健康,提高其非特异性免疫功能的作用。
1 甘露寡糖对鱼类生长性能的影响
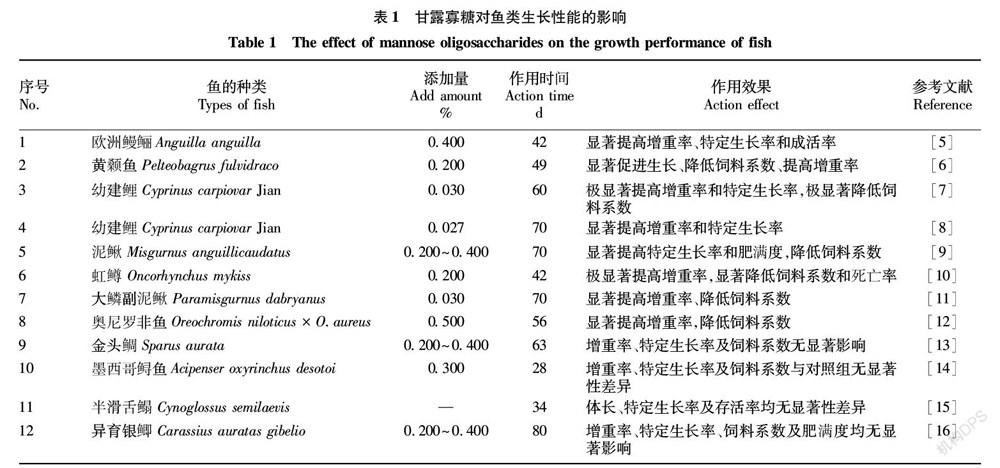
国内外关于甘露寡糖(MOS)对鱼类生长性能影响的研究结果尚存在差异,部分研究表明在鱼类日粮中添加甘露寡糖可以提高鱼类的生长性能,其作用方式主要通过提高鱼类体增重率、降低饲料系数等方面来实现(表1)。在对欧洲鳗鲡(Anguilla anguilla)[5]、黄颡鱼(Pelteobagrus fulvidraco)[6]、幼建鲤(Cyprinus carpiovar Jian)[7-8]、泥鳅(Misgurnus anguillicaudatus)[9]、虹鳟(Oncorhynchus mykiss)[10]及大鳞副泥鳅(Paramisgurnus dabryanus)[11]的研究中均发现,甘露寡糖能够通过提高增重率、特定生长率,降低饲料系数等来提高鱼类的生长性能。但在对奥尼罗非鱼(Oreochromis niloticus × O.aureus)[12]的研究中发现,甘露寡糖在鱼类饲料中的添加量有一个合适的范围,过量的添加可能会对鱼类的生长产生负面影响,原因可能是较高浓度的甘露寡糖促进肠道绒毛高度过度发育导致肠道阻塞,抑制鱼类对营养物质的消化吸收,从而对鱼类的生长性能产生了负面影响。日粮中添加甘露寡糖有助于动物的生长性能,其作用机制可能归因于改善了肠道健康,在一定的添加浓度范围内,甘露寡糖促进肠道绒毛高度、改善肠道菌群、提高肠道消化酶活力,导致肠道吸收面积增加、肠道菌群结构平衡、消化能力增强,营养物质吸收效率提高,从而促进鱼类的生长性能。
也有一些与之相反的研究,在金头鲷(Sparus aurata)[13]、墨西哥鲟鱼(Acipenser oxyrinchus desotoi)[14]、半滑舌鳎(Cynoglossus semilaevis)[15]及异育银鲫(Carassius auratas gibelio)[16]的研究上发现,基础饲料中添加甘露寡糖对其增重率、特定生长率、饲料系数等生长指标均无显著性影响。原因可能是试验鱼类种类或鱼体生长发育阶段不同,对试验结果产生了不同的影响;试验过程差异,试验者选择甘露寡糖的作用时间、添加量、养殖环境以及饲养周期的长短都有可能导致甘露寡糖对鱼类的生长作用产生差异。但从总体来看,甘露寡糖能够提高鱼类的生长性能。
2 甘露寡糖对鱼类肠道功能的影响
2.1 甘露寡糖对鱼类肠道菌群结构的影响
鱼类肠道内定植着数量庞大的微生物群落,主要分为有益微生物菌群(乳酸杆菌属、双歧杆菌属等)和有害微生物菌群(大肠杆菌、嗜水气单胞菌属等)[17],这些微生物菌群共同组成了鱼体肠道的微生物区系,直接影响着机体的营养和健康。大量研究结果表明,在鱼类日粮中添加甘露寡糖与鱼类肠道菌群平衡之间有着直接的关系。张红梅[18]在鲤鱼(Cyprinus carpio)的基础饲料中添加甘露寡糖,随着剂量的增加,鱼体肠道中的大肠杆菌数量减少与对照组有显著性差异(P<0.01),双歧杆菌数量增加与对照组相比差异极显著(P<0.01),乳酸杆菌数量增加与对照组相比差异不显著(P>0.05)。李云兰[8]在幼建鲤基础饲料中添加甘露寡糖,试验结果表明,当添加含量在180~270 mg/kg时,幼建鲤前、中和后肠大肠杆菌数量与对照组相比极显著降低(P<0.01);当甘露寡糖含量在270 mg/kg时,幼建鲤肠道中乳酸杆菌和双歧杆菌浓度相比对照组有显著性提高(P<0.05)。Torrecillas[19]在欧洲海鲈(Dicentrarchus labrax)的基础饲料中添加4 g/kg的甘露寡糖,8周试验后结果显示,甘露寡糖能够显著提高海鲈肠道中乳酸杆菌和双歧杆菌的数量,且显著降低了大肠杆菌和嗜水气单胞菌的数量。汪立平等[20]在奥尼罗非鱼饲料中添加自制酵母甘露寡糖,30 d试验结果表明在罗非鱼饲料中添加0.5%的甘露寡糖能够显著提高其肠道中的乳酸杆菌数量,降低肠道中大肠杆菌数量。苏健[21]在泥鳅的日粮中添加甘露寡糖,试验结果表明,当添加量为200和400 mg/kg时,甘露寡糖显著降低了泥鳅肠道中大肠杆菌的数量,且显著增加了肠道中乳酸杆菌和双歧杆菌的数量。甘露寡糖可以与有害细菌的受体竞争结合,降低有害细菌在肠道定值的几率,加速有害细菌从肠道中排出,同时这种竞争性排斥作用也有利于有益菌的生长,此外甘露寡糖还能够通过降低肠道pH值,抑制有害菌的生长[22]。因此,甘露寡糖对肠道菌群的调节主要在于促进肠道有益菌增殖抑制有害菌,优化肠道菌群结构,促进肠道微生物区系的平衡,从而提高动物的生长性能和免疫能力。
2.2 甘露寡糖对鱼类肠道形态结构的影响
肠道是鱼类容纳消化食物和运输吸收营养物质的主要场所,肠道健康是鱼类正常生长发育的重要保证[23]。肠道形态是最能直观反映魚类肠道健康的指标,肠道绒毛高度、肌层厚度与肠道的吸收能力相关,直接影响着鱼体的消化吸收营养物质的能力[24]。小肠绒毛的主要功能是吸收营养物质,小肠绒毛高度的变化决定了肠道有效吸收面积的大小,肠道皱襞高度宽度的增加也有利于营养物质的消化吸收,体现着肠道的健康状况。甘露寡糖能够改变肠道形态结构,可以增加猪[25]、兔[26]、鸡[27]等畜禽动物的小肠绒毛高度和宽度。在水产动物的研究中,胡凌豪等[28]在斜带石斑鱼(Epinephelus coioides)基础饲料中添加不同剂量的果寡糖,试验结果表明,果寡糖能够增加斜带石斑鱼的肠道绒毛高度和宽度,且0.1%试验组肠道的肌层厚度显著高于对照组(P<0.05);刘爱君等[12]在奥尼罗非鱼的基础饲料中添加甘露寡糖,对各组肠道做组织切片后发现,添加0.25%、0.50%和0.75%的甘露寡糖均能显著增加肠道绒毛高度、绒毛宽度和绒毛密度(P<0.05),且0.50%组的效果最显著;徐磊[16]在异育银鲫的日粮中添加甘露寡糖养殖80 d,120 mg/kg组的肠绒毛长度相比对照组提高了8.1%,但各试验组肠绒毛长度与肠直径的比值、皱襞间质宽、黏膜下层宽、肌层宽与对照组无显著性差异(P>0.05);Torrecillas等[19] 在欧洲海鲈的基础饲料中添加4 g/kg的甘露寡糖,试验结束后与对照组相比试验组鱼体肠道前肠黏膜的褶皱高度,宽度和褶皱表面积都有显著性增加(P<0.05),后肠总表面积增加(P<0.05),但后肠褶皱较对照组短(P<0.05);Dimitroglou等[29]在虹鳟鱼的基础饲料中添加0.2%的甘露寡糖,用光镜和电子显微镜观察前肠和后肠形态发现,与对照组相比试验组的肠道吸收面积增加,微绒毛长度和密度显著增加(P<0.05),但在仔鱼的研究上结果差异并不显著,可能的原因是甘露寡糖对不同生长发育阶段鱼体的影响有差异。Dimitroglou等[30]在白鲷(Diplodus sargus L.)的卤虫饲料中添加甘露寡糖,结果表明,与对照组相比添加甘露寡糖可将其微绒毛长度增加26%;在大菱鲆肠道健康(Scophthalmus maximus)的研究上,发现在基础饲料中添加甘露寡糖能改善肠道绒毛高度和黏膜皱襞,抑制微绒毛和黏膜皱襞的改变,改善大菱鲆肠道健康状况[31]。甘露寡糖对鱼类肠道形态结构产生有益影响,肠道结构的改善导致营养物质消化率和饲料利用率的提高,从而促进鱼类的生长性能。Pryor等[14]在墨西哥鲟鱼饲料中添加0.3%的甘露寡糖,26 d试验结果表明MOS对其肠道绒毛高度、宽度和密度均无显著性影响;Salem等[32]在欧洲黑鲈日粮中添加的0.2%、0.4%甘露寡糖对肠道绒毛长度没有影响。以上差异可能是由于甘露寡糖的添加量不同、鱼的种类不同、鱼类机体肠道内的肠道微生物群不同或者试验方法不同等。
2.3 甘露寡糖对鱼类肠道消化酶的影响
动物机体的消化能力除了与肠道生态环境、形态结构相关联外还与肠道中消化酶的活力密切联系。鱼类肠道消化酶是反映机体消化能力和吸收营养成分的重要指标。研究表明功能性寡糖能够促进肠道上皮细胞生长,提高内源酶活性,促进鱼类生长发育。杨敏等[5]在欧洲鳗鲡的饲料中添加不同含量的甘露寡糖,42 d试验后发现,甘露寡糖能够提高欧洲鳗鲡肠道胰蛋白酶、淀粉酶和脂肪酶活性,0.4%组胰蛋白酶、脂肪酶和淀粉酶活性与对照组有显著性差异(P<0.05);强俊等[33]在奥尼罗非鱼幼鱼饲料中添加不同水平的低聚木糖(XOS),结果表明添加低聚木糖组提高了奥尼罗非鱼幼鱼肠道淀粉酶和脂肪酶活力,分别在0.030%和0.045%最高,且各组之间有显著性差异(P<0.05);李洋[7]在幼建鲤基础饲料中添加甘露寡糖,试验结果表明,添加量为300 mg/kg时糜蛋白酶和肠淀粉酶极显著高于对照组(P<0.05),肠脂肪酶显著高于对照组(P<0.05),但胰蛋白酶与对照组差异不显著(P>0.05);此外,Safai等[34]在小龙虾日粮中分别添加甘露寡糖和低聚果糖(FOS)及其组合,结果表明,MOS和FOS及组合添加对其生长性能、消化酶活力等均有显著性提高,但MOS和FOS的组合添加比起单独添加作用效果更好。
3 甘露寡糖对鱼类免疫功能的影响
动物机体的免疫系统包括特异性免疫和非特异性免疫,非特异性免疫系统是阻止病原菌入侵机体的第一道防线,在鱼类机体中发挥着重要作用,对其生长和抵御病原入侵有着重要的影响[35]。当病原微生物和异物入侵时,非特异性免疫通过物理屏障、吞噬作用、溶菌作用和凝集作用等清除病原菌和异物[36]。鱼类非特异性免疫能力的提高主要表现在抑菌类物质的产生,吞噬细胞吞噬能力及相关免疫酶活力等方面的提高。
3.1 甘露寡糖对鱼类免疫酶活力的影响
溶菌酶是非特异性免疫系统的重要组成,是存在于鱼类体表黏液、血清和巨噬细胞中的一种水解酶,在动物机体排斥异物攻击时发挥着重要作用;酸性磷酸酶存在于巨噬細胞溶酶体,是巨噬细胞内最具代表性的水解酶之一,能够快速对异物识别、吞噬和清除达到预防感染的目的[37];碱性磷酸酶广泛存在于动物血液和各种器官内,能够通过改变病原体表面结构,增强机体对病原体的识别和吞噬能力[38]。因此其酶活力可作为免疫指标来间接反映鱼类非特异免疫功能。大量研究表明,甘露寡糖可以增强鱼类免疫酶活力,Lu等[39]在草鱼(Ctenopharyngodon idella)基础饲料中添加甘露寡糖,研究结果表明MOS可以增强其溶菌酶和磷酸酶活力;张红梅等[40]在鲤鱼的基础饲料中添加不同水平的甘露寡糖,结果显示随着添加量的增加血清中溶菌酶活力呈增长趋势,其中0.3%添加组与对照组有极显著差异;刘爱君等[41]报道在奥尼罗非鱼基础饲料中添加甘露寡糖能显著提高其血清溶菌酶活力和血清碱性磷酸酶活力;Ayce Genc等[42]在罗非鱼(Oreochromis mossambicus)基础饲料中添加3 g/kg的甘露寡糖显著提高了其血清溶菌酶活力;王锐等[43]在异育银鲫幼鱼饲料中添加半乳甘露寡糖,试验结果表明甘露寡糖显著提高了异育银鲫幼鱼血清溶菌酶的活力。此外,Meng等[44]在甘露寡糖对皱纹盘鲍(Haliotis discus hannai Ino)幼鲍的研究上也发现,甘露寡糖能显著提高其肝胰脏和血清中碱性磷酸酶、酸性磷酸酶和溶菌酶活力。因此,甘露寡糖可以提高鱼类的免疫酶活性,进而增强其非特异性免疫功能。甘露寡糖能够优化肠道微生物区系促进有益菌的增殖,而双歧杆菌和乳酸杆菌的增殖提高了机体抗体水平,促进了巨噬细胞的吞噬能力[45],从而提高相关免疫酶活性对机体非特异性免疫能力产生有益影响。
3.2 甘露寡糖对鱼类红细胞数量的影响
红细胞是鱼类机体中重要的免疫细胞,能够影响机体的吞噬能力,在鱼类的非特异性免疫防御中发挥着重要作用[46]。李云兰[7]发现甘露寡糖极显著的影响了幼建鲤血液红细胞数量(P<0.01),且当甘露寡糖添加量在619 mg/kg时,幼建鲤血液红细胞数量最多增加58.23%;Andrews等[47]在南亚野鳞幼鱼(Labeo rohita)饲料中添加1%的甘露寡糖,结果表明其血液红细胞数量相比对照组提高了5.3%;李洋[7]报道添加甘露寡糖极显著增加了幼建鲤血液红细胞数量。此外,Rodriguez-Estrada等[48]在虹鳟日粮中分别添加粪肠球菌(EF)和甘露寡糖(MOS)及其组合,结果表明EF和MOS以及联合使用显著提高了虹鳟的红细胞压积值,但分析结果表明EF和MOS的联合添加对虹鳟的红细胞压积值、粘液重量等免疫指标的提升达到最佳水平。上述研究结果说明,甘露寡糖能够通过提高鱼类血液红细胞数量来增强机体非特异免疫能力。
3.3 甘露寡糖对鱼类白细胞吞噬率的影响
水生动物体内的吞噬细胞主要包括中性粒细胞和单核巨噬细胞,是白细胞的主要成分。鱼类血液中白细胞吞噬能力可以反映出机体的免疫状态,白细胞吞噬能力一般用白细胞吞噬率和吞噬指数来表示。鱼类机体吞噬细胞对病原微生物的吞噬和杀灭作用,是其非特异性免疫的一个重要方面[49]。王锐等[43]研究发现半乳甘露寡糖组的异育银鲫白细胞数目、吞噬细胞百分比均显著高于对照组(P<0.05),甘露寡糖组的吞噬指数相比对照组显著提高(P<0.05),且0.2%添加组最高;马志红等[50]在鲤基础饲料中添加0.1%的甘露寡糖,结果表明试验组相比对照组白细胞吞噬率、吞噬指数均显著增加(P<0.01);李洋[7]研究发现甘露寡糖极显著提高了幼建鲤血液白细胞吞噬率(P<0.01);Torrecillas等[51]发现甘露寡糖可以显著提高欧洲海鲈的白细胞数量和白细胞吞噬率;Yoshida等[52]发现在大西洋鲶(Clarias gariepinus)基础饲料中添加甘露寡糖能够显著提高其嗜中性粒细胞的吞噬活性。因此,甘露寡糖能够通过提高鱼类的白细胞吞噬率从而提高其非特异性免疫功能。Spring[53]研究表明吞噬细胞吞噬能力提高的原因可能是甘露寡糖促进了辅助性T-淋巴细胞的活性,甘露寡糖能够增强辅助性T-淋巴细胞分泌白细胞介素-2(IL-2)和γ-干扰素(IFN-γ)的能力,从而对机体的非特性免疫能力产生积极的影响。
4 小结和展望
作为一种绿色饲料添加剂,甘露寡糖能够提高动物机体的生长性能,促进肠道发育,优化肠道菌群结构并增强其非特异免疫功能,能够广泛应用在动物饲料行业,是抗生素的良好替代品。但甘露寡糖在水产养殖中的应用仍有一些问题有待解决:
(1)关于甘露寡糖对水产动物的研究还不够全面,未来仍然需要探索甘露寡糖在更多经济水产动物饲料中的适宜添加量、不同生长发育阶段的使用方式、不同种类甘露寡糖的使用效果等,从而使其在水产养殖中得到更深入广泛地应用。
(2)甘露寡糖与其他饲料添加剂的联合使用。已有部分试验表明甘露寡糖与其他添加剂的联合使用比起单独添加对水产动物的作用效果似乎更好,因此,未来应该致力于功能性寡糖之间或者与其他营养素之间相关性的研究,以期更加有效率地使用饲料添加剂,从而实现高效率、低成本、无污染的水产养殖。
参考文献
[1]鲍延安,邢淑芳,徐庆龙.甘露寡糖对荷斯坦奶牛产奶量及乳常规的影响[J].饲料研究,2009(2):57-60.
[2] 熊阿玲.日粮添加甘露寡糖对肉鸡生长性能及天然免疫相关基因表达的影响研究[D].南昌:江西农业大学,2014.
[3] CHACHER M F A,KAMRAN Z,AHSAN U,et al.Use of mannan oligosaccharide in broiler diets:An overview of underlying mechanisms[J].World′s poultry science journal,2017,73(4):831-844.
[4] AGAZZI A,PERRICONE V,ZORINI F O,et al.Dietary mannan oligosaccharides modulate gut inflammatory response and improve duodenal villi height in post-weaning piglets improving feed efficiency[J].Animals,2020,10(8):1-14.
[5] 杨敏,黎中宝,卢静,等.甘露寡糖对欧洲鳗鲡(Anguilla anguilla)生长、消化酶活性及非特异性免疫的影响[J].中国渔业质量与标准,2016,6(6):40-46.
[6] 于艳梅,吴志新,陈孝煊,等.魔芋甘露寡糖对黄颡鱼非特异性免疫功能及生长的影响[J].华中农业大学学报,2010,29(3):351-355.
[7] 李洋.甘露寡糖对幼建鲤生长性能和非特异性免疫的影响[D].雅安:四川农业大学,2011.
[8] 李云兰.甘露寡糖对幼建鲤(Cyprinus carpio Var.Jian)肠道菌群和免疫功能的影响[D].雅安:四川农业大学,2004.
[9] 杜宗君,苏健,巫红萍,等.甘露寡糖对泥鳅生产性能、肠道菌群和非特异性免疫功能的影响[J].四川农业大学学报,2019,37(1):129-133.
[10] STAYKOV Y,SPRING P,DENEV S,et al.Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss)[J].Aquaculture international,2007,15(2):153-161.
[11] XU B,WU S J,HAN Q.Modulation of the growth performance and innate immunity of loaches (Paramisgurnus dabryanus) upon dietary mannan oligosaccharides[J].3 Biotech,2021,11(3):1-7.
[12] 刘爱君,冷向军,李小勤,等.甘露寡糖对奥尼罗非鱼(Oreochromis niloticus×O.aureus)生长、肠道结构和非特异性免疫的影响[J].浙江大学学报(农业与生命科学版),2009,35(3):329-336.
[13] DIMITROGLOU A,MERRIFIELD D L,SPRING P,et al.Effects of mannan oligosaccharide (MOS) supplementation on growth performance,feed utilisation,intestinal histology and gut microbiota of gilthead sea bream (Sparus aurata)[J].Aquaculture,2010,300(1/2/3/4):182-188.
[14] PRYOR G S,ROYES J B,CHAPMAN F A,et al.Mannanoligosaccharides in fish nutrition: Effects of dietary supplementation on growth and gastrointestinal villi structure in gulf of Mexico sturgeon[J].North American journal of aquaculture,2003,65(2):106-111.
[15] 于朝磊,常青,吕云云.甘露寡糖对半滑舌鳎(Cynoglossus semilaevis Günther)稚魚生长、肠道发育和非特异性免疫水平的影响[J].渔业科学进展,2014,35(6):53-59.
[16] 徐磊.甘露寡糖对异育银鲫生长性能及抗嗜水气单胞菌感染的研究[D].南京:南京农业大学,2010.
[17] LLEWELLYN M S,BOUTIN S,HOSEINIFAR S H,et al.Teleost microbiomes: The state of the art in their characterization,manipulation and importance in aquaculture and fisheries[J].Frontiers in microbiology,2014,5:1-17.
[18] 张红梅.甘露寡聚糖对鲤鱼生产性能及对鱼体各项生物学指标的影响[D].保定:河北农业大学,2003.
[19] TORRECILLAS S,MAKOL A,BENTEZ-SANTANA T,et al.Reduced gut bacterial translocation in European sea bass (Dicentrarchus labrax) fed mannan oligosaccharides (MOS)[J].Fish & shellfish immunology,2011,30(2):674-681.
[20] 汪立平,马相杰,冷向军,等.甘露寡糖对罗非鱼肠道菌群的影响[J].食品工业科技,2010,31(10):142-145.
[21] 苏健.甘露寡糖对泥鳅生产性能、肠道菌群和非特异性免疫功能的影响[D].雅安:四川农业大学,2013.
[22] 李凤娜,王继成.寡糖对肠道菌群的调节及常用研究方法[J].兽药与饲料添加剂,2005,10(4):23-25.
[23] 涂永锋,宋代军.鱼类肠道组织结构及其功能适应性[J].江西饲料,2004(4):16-19.
[24] 李霞.水产动物组织胚胎学[M].北京:中国农业出版社,2006.
[25] 黄俊文,林映才,冯定远,等.纳豆菌、甘露寡糖对仔猪肠道pH、微生物区系及肠黏膜形态的影响[J].畜牧兽医学报,2005,36(10):1021-1027.
[26] MOURO J L,PINHEIRO V,ALVES A,et al.Effect of mannan oligosaccharides on the performance,intestinal morphology and cecal fermentation of fattening rabbits[J].Animal feed science & technology,2006,126(1/2):107-120.
[27] IJI P A,SAKI A A,TIVEY D R.Intestinal structure and function of broiler chickens on diets supplemented with a mannan oligosaccharide[J].Journal of the science of food & agriculture,2001,81(12):1186-1192.
[28] 胡凌豪,杨红玲,赵芸,等.果寡糖对斜带石斑鱼免疫功能和肠道形态的影响[J].水产科学,2019,38(5):589-594.
[29] DIMITROGLOU A,MERRIFIELD D L,MOATE R,et al.Dietary mannan oligosaccharide supplementation modulates intestinal microbial ecology and improves gut morphology of rainbow trout,Oncorhynchus mykiss(Walbaum)[J].Journal of animal science,2009,87(10):3226-3234.
[30] DIMITROGLOU A,DAVIES S J,SWEETMAN J,et al.Dietary supplementation of mannan oligosaccharide on white sea bream (Diplodus sargus L.) larvae:Effects on development,gut morphology and salinity tolerance[J].Aquaculture Research,2010,41(9):e245-e251.
[31] BAI N,GU M,XU X J,et al.Protective effects of mannan oligosaccharides on turbot Scophthalmus maximus suffering from soy enteropathy[J].Aquaculture,2017,476:141-151.
[32] SALEM M,GABER M M,ZAKI M A D,et al.Effects of dietary mannan oligosaccharides on growth,body composition and intestine of the sea bass ( Dicentrarchus labrax L.)[J].Aquaculture research,2016,47(11):3516-3525.
[33] 強俊,王辉,李瑞伟,等.低聚木糖对奥尼罗非鱼幼鱼生长、体成分和消化酶活力的影响[J].淡水渔业,2009,39(6):63-68.
[34] SAFARI O,SHAHSAVANI D,PAOLUCCI M,et al.Single or combined effects of fructo-and mannan oligosaccharide supplements on the growth performance,nutrient digestibility,immune responses and stress resistance of juvenile narrow clawed crayfish,Astacus leptodactylus leptodactylus Eschscholtz,1823[J].Aquaculture,2014,432:192-203.
[35] 钱云霞,王国良,邵健忠.鱼类的非特异性免疫调节[J].宁波大学学报(理工版),2000,13(1):95-99.
[36] 慈麗宁,刘波,谢骏,等.影响水生动物免疫机能的因素(综述)[J].江苏农业科学,2012,40(3):205-210.
[37] CHENG T C.The role of lysosomal hydrolases in molluscan cellular response to immunologic challenge[M]//BULLA L A JR,CHENG T C.Invertebrate models for biomedical research.New York:Plenum Press,1978.
[38] 牟海津,江晓路,刘树青,等.免疫多糖对栉孔扇贝酸性磷酸酶、碱性磷酸酶和超氧化物歧化酶活性的影响[J].青岛海洋大学学报(自然科学版),1999,29(3):463-468.
[39] LU Z Y,JIANG W D,WU P,et al.Mannan oligosaccharides supplementation enhanced head-kidney and spleen immune function in on-growing grass carp (Ctenopharyngodon idella)[J].Fish & shellfish immunology,2020,106(4):596-608.
[40] 张红梅,姜会民.酵母甘露寡糖对鲤鱼非特异性免疫的影响[J].饲料研究,2006(10):25-27.
[41] 刘爱君,冷向军,李小勤,等.黄霉素和甘露寡糖对奥尼罗非鱼的生长性能及血清非特异性免疫的影响[J].中国饲料,2009(3):29-32.
[42] AYCE GENC M,YILMAZ E,GENC E,et al.Effects of dietary mannan oligosaccharides (MOS) on growth,body composition,and intestine and liver histology of the hybrid tilapia (Oreochromis niloticus×O.aureus)[J].The Israeli journal of aquaculture-bamidgeh,2007,59(1):10-16.
[43] 王锐,刘军,刘辉宇,等.半乳甘露寡糖对异育银鲫幼鱼生长和非特异性免疫的影响[J].上海水产大学学报,2008,17(4):502-506.
[44] MENG X X,YANG X Y,LIN G,et al.Mannan oligosaccharide increases the growth performance,immunity and resistance capability against Vibro Parahemolyticus in juvenile abalone Haliotis discus hannai Ino[J].Fish & shellfish immunology,2019,94:654-660.
[45] 唐凌,邝声耀,冯琳,等.甘露寡糖对水生动物肠道菌群和免疫功能的影响[J].中国畜牧兽医,2008,35(6):21-25.
[46] 蔡完其,轩兴荣.红鲤4群体间红细胞免疫功能及其差异[J].中国水产科学,2003(2):133-136.
[47] ANDREWS S R,SAHU N P,PAL A K,et al.Haematological modulation and growth of Labeo rohita fingerlings:Effect of dietary mannan oligosaccharide,yeast extract,protein hydrolysate and chlorella[J].Aquaculture research,2009,41(1):61-69.
[48] RODRIGUEZ-ESTRADA U,SATOH S,HAGA Y,et al.Effects of inactivated Enterococcus faecalis and mannan oligosaccharide and their combination on growth,immunity,and disease protection in Rainbow trout[J].North American journal of aquaculture,2013,75(3):416-428.
[49] 肖克宇.水产动物免疫与应用[M].北京:科学出版社,2007.
[50] 马志红,晋晶,刘洪梅,等.甘露寡糖对鲤鱼生长性能和免疫功能的影响[J].安徽农业科学,2010,38(3):1291-1292,1296.
[51] TORRECILLAS S,MAKOL A,CABALLERO M J,et al.Immune stimulation and improved infection resistance in European sea bass (Dicentrarchus labrax) fed mannan oligosaccharides[J].Fish & shellfish immunology,2007,23(5):969-981.
[52] YOSHIDA T,KRUGER R,INGLIS V.Augmentation of non-specific protection in African catfish,Clarias gariepinus (Burchell),by the long‐term oral administration of immunostimulants[J].Journal of fish diseases,1995,18(2):195-198.
[53] SPRING P,WENK C,DAWSON K A,et al.The effects of dietary mannaoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of salmonella-challenged broiler chicks[J].Poultry science,2000,79(2):205-211.
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
儿童时代·幸福宝宝(2020年9期)2020-09-08
新高考(英语进阶)(2017年10期)2017-12-23
探索科学(2017年5期)2017-06-21
湖北农业科学(2016年18期)2016-12-08
河北渔业(2016年10期)2016-11-22
金色少年(奇趣科普)(2016年8期)2016-09-21
江苏农业科学(2015年8期)2015-09-10
河北渔业(2015年2期)2015-03-26
