
饲粮添加色氨酸对热应激肉鸡肝脏、肠道抗氧化能力和炎症反应的影响
2022-07-15 00:49欧阳经鑫李秋粉郭帅鹏刘三凤黎观红江西农业大学江西省动物营养重点实验室江西南昌330045江西农业大学动物科学技术学院江西南昌330045
中国兽医学报 2022年6期
欧阳经鑫,李秋粉,周 华,郑 俊,郭帅鹏,刘三凤,黎观红* (1.江西农业大学 江西省动物营养重点实验室,江西 南昌 330045;.江西农业大学 动物科学技术学院,江西 南昌 330045)
随着全球气候变暖以及集约化养殖的普及,加之中国南方地区高温高湿的气候特征,热应激是畜牧业发展需面临的严峻挑战。当环境温度超过机体的等热区上限,机体不能维持产热和散热的稳态,就会发生热应激[1]。而家禽因其代谢速率快、体温偏高、被覆羽毛、缺乏汗腺等生理特性,更容易受到热应激[2]。热应激对肉鸡的影响涉及许多方面,包括使畜禽体温度升高,采食量降低,代谢异常,氧化还原稳态失衡,免疫机能、屏障功能下降等现象,进而导致机体组织器官的损伤甚至是死亡[3-6]。当机体组织受损后,受损细胞释放内源性损伤相关分子模式(DAMPs)并激活下游信号通路,诱导炎症因子的分泌而引发炎症反应[7-9]。研究表明活性氧,特别是来自线粒体的活性氧,可以激活NOD样受体热蛋白结构域相关蛋白3(NLRP3)炎症小体,增加白细胞介素(IL)-1β、IL-18的释放[10-12]。张宏男等[13]研究发现热应激可以激活NLRP3炎症小体,敲除NLRP3基因可以减轻全身性炎症反应,维持肠道屏障的完整性,减轻组织的损伤程度。因此,NLRP3炎症小体的激活可能参与了热应激导致的炎症反应过程。
色氨酸(Trp)是畜禽的必需氨基酸,也是家禽饲粮中的限制性氨基酸[14]。色氨酸及其代谢产物在动物采食量、行为、应激和免疫反应等方面发挥着重要的调节作用[15]。岳云双[16]研究发现,饲粮添加色氨酸可以提高应激条件下肉鸡的生产性能,抑制应激反应引起的肠道促炎细胞因子分泌,维持肠道正常屏障功能。褪黑素是色氨酸的代谢产物之一,被认为是一种有效的自由基清除剂和氧化还原活性酶的调节剂[17]。尹华祺[18]研究发现,褪黑素可以增加热应激小鼠血液中白细胞、单核细胞、粒细胞等免疫细胞数量,调节免疫功能,提高超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)等抗氧化酶活性,降低丙二醛(MDA)的含量,有效缓解热应激导致的组织损伤。因此,本研究以AA肉鸡为试验对象,探讨热应激对肉鸡肝脏和肠道抗氧化能力及炎症反应的影响和饲粮添加色氨酸的调节作用,为色氨酸缓解动物应激损伤提供进一步的试验依据。
1 材料与方法
1.1 试验设计选取健康、体质量接近的180只18日龄雌性AA肉鸡,随机分为3个处理组,每组6个重复,每个重复10只。适应性饲养至21日龄开始进行温度控制,试验期为21 d。适温组(TN)肉鸡饲养于正常温度(23±1)℃,饲喂基础饲粮;热应激组(HS)肉鸡饲养于高温环境(34±1)℃、8 h/d,饲喂基础饲粮;热应激+0.18%色氨酸组(HS+0.18% Trp)肉鸡饲养于高温环境(34±1)℃、8 h/d,饲喂基础饲粮+0.18%色氨酸。本试验所用的L-色氨酸购自韩国希杰食品有限公司,纯度>98%。
1.2 试验日粮与动物饲养管理本试验采用玉米-豆粕型基础饲粮,参考NRC(1994)推荐的营养水平进行配制,基础日粮组成及营养水平见表1。通过调节谷氨酸添加量使各组饲粮保持等氮等能水平。试验在江西农业大学试验基地的试验鸡房进行。试验鸡采用4层笼笼养。试验开始前对鸡舍及周围环境进行充分冲洗和严格消毒,鸡只按正常免疫程序进行免疫。试验期热应激从21日龄开始至42日龄结束。HS组温度控制如下:每天从9:00-17:00 维持(34±1)℃ 8 h,共21 d。试验期间鸡只自由采食和饮水。
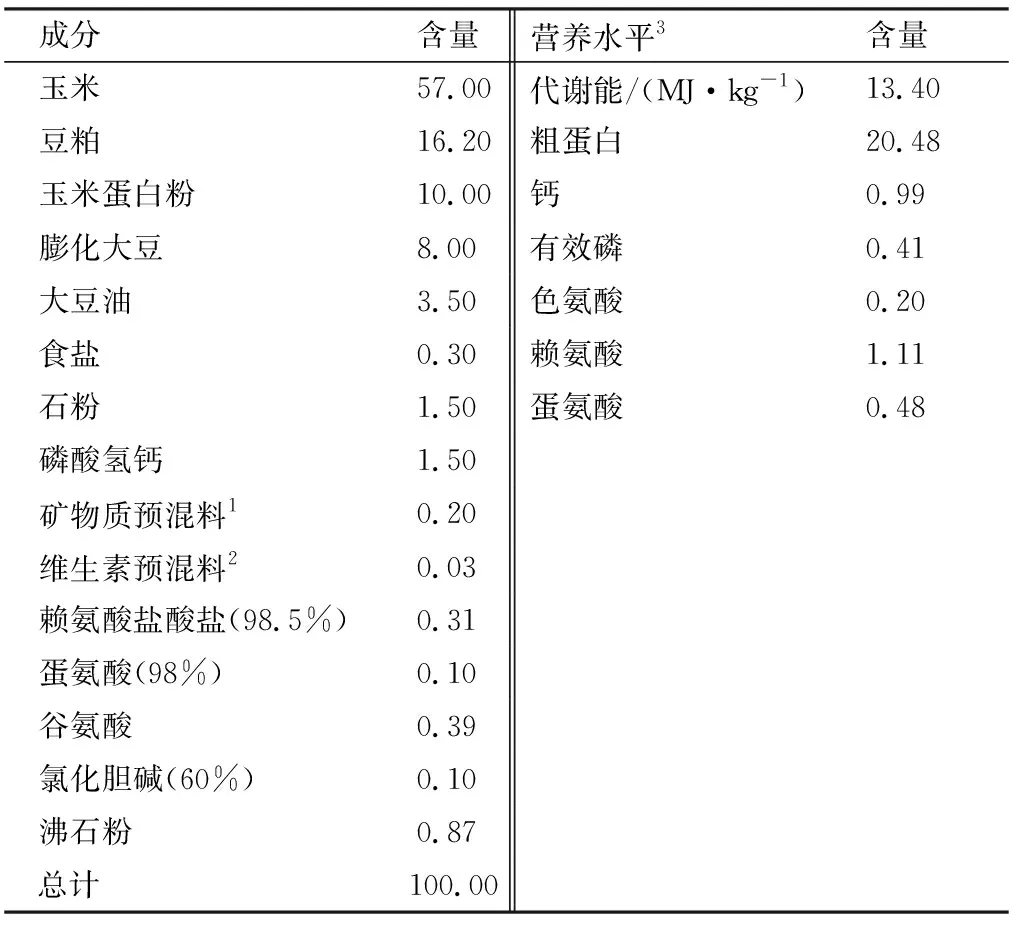
表1 基础饲粮组成及营养水平(风干基础) %
1.3 样品采集与制备于42日龄时,肉鸡提前禁食12 h,期间自由饮水。从每个重复中随机选取 1 只鸡,断颈处死,剖开腹腔,取肝脏、空肠和回肠,用PBS冲洗干净肠道内容物,吸水纸吸干后,刮取空肠、回肠黏膜,置于无菌无酶冻存管中,液氮速冻,-80℃ 保存用于后续分析。
1.4 测定指标与方法
1.4.1肝脏、肠道黏膜抗氧化指标的测定 用生理盐水制备10%的组织匀浆,离心取上清进行试验检测,采用BCA法测定上清液的蛋白浓度。采用南京建成生物工程研究所的试剂盒,严格按照说明书操作测定肝脏、肠道黏膜SOD、过氧化氢酶(CAT)、GSH-Px的活力、总抗氧化能力(T-AOC)和MDA的含量。
1.4.2肝脏细胞因子含量的测定 用PBS制备10%的肝脏组织匀浆,5 000×g离心10 min取上清进行检测,采用BCA法测定上清液的蛋白浓度。采用酶联免疫吸附测定(ELISA)试剂盒(上海酶联生物科技有限公司),检测肝脏中IL-1β、IL-6、IL-12、IL-18和肿瘤坏死因子α(TNF-α)的含量。检测方法严格按照说明书进行。
1.4.3实时荧光定量PCR 使用TransZol Up(北京全式金生物技术有限公司)裂解样品,提取肉鸡肝脏和肠道黏膜总RNA,具体操作按说明书进行。RNA提取完成后,用微量紫外分光光度计(NanoDrop ND-1000)测定RNA样品的纯度和浓度,保证D260 nm/D280 nm比值均在1.8~2.0之间,使用EasyScript®One-Step gDNA Removal and cDNA Synthesis SuperMix反转录试剂盒(北京全式金生物技术有限公司),按说明加入相应体系,将样品RNA反转录成 cDNA,于-20℃保存备用。根据PerfectStartTMGreen qPCR SuperMix试剂盒进行实时荧光定量反应,采用以下热循环条件:94℃预变性30 s;94℃变性5 s,在57~60℃退火延伸30 s,共循环42次。目的基因和内参基因β-actin的引物序列见表2,并由华大基因设计合成。以β-actin为内参基因,采用2-ΔΔCt法计算目的基因的相对表达量。

表2 荧光定量PCR引物序列

2 结果
2.1 饲粮添加色氨酸对热应激肉鸡肝脏和肠道黏膜抗氧化能力的影响由表3可知,与TN组相比,HS组肉鸡肝脏GSH-Px、回肠黏膜CAT活性和T-AOC显著降低(P<0.05),肝脏CAT、回肠黏膜SOD活性极显著降低(P<0.01);与HS组相比,HS+0.18%Trp组肝脏、空肠黏膜T-AOC显著升高(P<0.05),肝脏CAT、空肠和回肠黏膜SOD活性极显著升高(P<0.01),然而各组间的MDA含量无显著性差异(P>0.05)。

表3 饲粮添加色氨酸对热应激肉鸡肝脏和肠道黏膜抗氧化能力的影响
2.2 饲粮添加色氨酸对热应激肉鸡肝脏炎症细胞因子含量的影响由表4可知,与TN组相比,热应激显著升高肝脏IL-6和TNF-α的含量(P<0.05),极显著提高肝脏IL-1β和IL-18的含量(P<0.01)。与HS组相比,添加0.18% Trp显著降低热应激肉鸡肝脏IL-1β和TNF-α的含量(P<0.05),极显著降低热应激肉鸡肝脏IL-18的含量(P<0.01)。

表4 饲粮添加色氨酸对热应激肉鸡肝脏炎症细胞因子含量的影响 ng/g
2.3 饲粮添加色氨酸对热应激肉鸡肝脏、肠道黏膜炎症小体相关基因相对表达量的影响由表5可知,与TN组相比,热应激显著升高肝脏IL-1β和IL-18以及空肠黏膜IL-12A、IL-12B和IL-18的mRNA相对表达量(P<0.05),并极显著提高空肠黏膜NLRP3、IL-1β和TNF-α的mRNA相对表达量(P<0.01)。与TN组相比,热应激显著升高回肠黏膜NLRP3的mRNA相对表达(P<0.05),并极显著升高回肠黏膜Caspase-1、IL-1β和TNF-α的mRNA相对表达量(P<0.01)。与HS组相比,饲粮添加色氨酸显著降低热应激肉鸡肝脏TNF-α以及空肠黏膜IL-12A和IL-12B的mRNA相对表(P<0.05),并极显著降低热应激肉鸡空肠黏膜NLRP3、IL-18和TNF-α的mRNA相对表达量(P<0.01)。与HS组相比,饲粮添加色氨酸极显著降低热应激肉鸡回肠黏膜NLRP3、Caspase-1、IL-1β和TNF-α的mRNA相对表达量(P<0.01)。

表5 饲粮添加色氨酸对热应激肉鸡肝脏、肠道黏膜炎症小体相关基因相对表达量的影响
3 讨论
热应激是氧化应激的重要诱导因子[6],使机体内有毒性的活性氧自由基过度积累,导致机体内氧化和抗氧化系统之间的不平衡,造成核酸、蛋白质、脂质等生物大分子的氧化损伤[19]。机体有自身抗氧化防御系统以维持机体自由基的产生和清除处于动态平衡,其中SOD、GSH-Px和CAT是组成抗氧化系统的第一道防线,SOD能够将O2-歧化成为H2O2,然后GSH-Px和CAT进一步将H2O2分解为H2O[20]。T-AOC能够反映机体防御体系的总抗氧化能力。MDA是多不饱和脂肪酸过氧化的最终产物,其含量能够反映机体脂质过氧化的程度[21]。本试验研究表明,与TN组相比,热应激显著降低SOD、GSH-Px、CAT抗氧化酶的活性及T-AOC的含量。这一结果与前人在肉鸡、绵羊上的研究结果一致[22-24],表明长期的热应激会破坏机体组织氧化-还原平衡,降低机体的抗氧化能力。色氨酸是血清素和褪黑素的重要前体,在维持肉鸡采食量、生长性能、免疫反应和抗应激等方面发挥着重要作用[25-26]。MUND等[27]研究发现,高于NRC推荐水平的色氨酸可提高肉鸡的生长性能、免疫反应和抗氧化状态。LIU等[28]研究发现,饲粮添加0.15%色氨酸能够提高断奶仔猪空肠黏膜SOD、GSH-Px和CAT活性,降低MDA含量,缓解由敌草快引起的肠道氧化应激损伤。在本研究中,饲粮添加色氨酸提高了SOD、CAT的活力及T-AOC的含量,这一结果与前人研究结果一致。以上结果表明,饲粮中添加色氨酸能够提高机体的抗氧化能力,清除过度产生的有毒性活性氧自由基,缓解热应激引起的肉鸡组织氧化应激损伤。
炎症是一种机体对抗外界有害刺激或病原体的自然防御机制[29]。适度的炎症反应可以帮助机体对抗病原体的入侵,清除坏死的细胞,促进机体正常功能的恢复。然而过度的炎症反应反而会加重损伤[30]。氧化应激造成组织细胞损伤并释放损伤相关模式分子,并被机体组织相应的模式识别受体识别进而激活下游信号通路,诱导炎症因子的表达,引起组织细胞的不可逆损伤[31]。TNF-α、IL-1β、IL-6是炎症反应中主要的炎症因子,少量分泌能够与免疫细胞上的受体结合发挥免疫调控作用,但随其分泌量增加,炎症反应被加剧而导致机体组织损伤[23]。IL-12能激活和诱导自然杀伤细胞的增殖、细胞毒性和细胞因子的产生[32]。DENG等[33]研究表明,34℃高温环境下持续热应激提高肉鸡血清TNF-α、IL-6和皮质酮的含量。TNF-α等促炎症因子的过量产生可导致包括肠道在内的器官出血和坏死[34]。李秋粉[35]研究报道,21 d的热应激显著升高肉鸡血清促炎因子TNF-α、IL-6的含量。本研究发现,与TN组相比,热应激升高肝脏炎症因子IL-1β、IL-18、IL-6、TNF-α的含量,这与前人的研究结果一致,同时也说明热应激能够促进炎症因子的分泌,引发机体炎症反应。NLRP3炎症小体是研究最为广泛及深入的炎症小体,并与多种人类自身炎症和自身免疫疾病有关[36]。炎症小体是由NOD样受体蛋白3(NLRP3)、细胞凋亡相关斑点样蛋白(ASC)以及半胱天冬酶前体(pro-Caspase-1)组成的细胞内多蛋白复合物,通过独特的模式识别受体响应多种外源性(包括脂多糖、细菌鞭毛等)和内源性危险信号(包括活性氧、K+外流、Ca2+内流和溶酶体损伤等)的刺激,招募和激活促炎蛋白酶Caspase-1,活化的Caspase-1可将pro-IL-1β和pro-IL-18切割成成熟的生物活性形式释放[13,36-37]。王伟等[38]研究表明,热应激可导致肝NLRP3炎症小体的激活,促进下游级联反应中Caspase-1、IL-1β、IL-6的表达上调,诱导肝脏炎症损伤,而抑制NLRP3表达水平可有效拮抗下游促炎因子的表达。在本研究中,热应激升高肉鸡NLRP3、Caspase-1及促炎细胞因子的mRNA相对表达量,此结果表明,NLRP3炎症小体激活下游级联反应共同参与热应激诱导的炎症反应,这与前人的研究结果一致[38]。本研究中,饲粮添加色氨酸显著降低热应激肉鸡肝脏炎症细胞因子及NLRP3信号通路的mRNA表达。YUE等[39]研究发现,慢性应激导致IL-1β、IL-6和TNF-α等促炎细胞因子mRNA水平升高以及抗炎细胞因子IL-10 mRNA表达量下降,而饲粮添加色氨酸能部分逆转以上变化,这与本试验结果基本一致。由此可见,色氨酸可能通过抑制NLRP3信号通路的表达,抑制机体的促炎反应,以此缓解由热应激引起的机体炎症损伤。
综上所述,长期热应激会导致肉鸡肝脏和肠道产生氧化应激,降低组织的抗氧化能力,同时诱发肝脏和肠道炎症反应。饲粮添加色氨酸能够缓解热应激引起的氧化应激损伤,通过抑制NLRP3炎症小体及下游级联反应,减轻炎症反应。
猜你喜欢
中国畜牧杂志(2022年6期)2022-06-13
中国畜牧杂志(2022年5期)2022-05-16
家禽科学(2020年3期)2020-05-13
现代畜牧科技(2019年5期)2019-07-08
现代畜牧科技(2018年7期)2018-10-21
科学种养(2018年7期)2018-09-13
国外畜牧学·猪与禽(2018年6期)2018-05-14
恋爱婚姻家庭·青春(2018年1期)2018-01-24
家庭医药(2016年1期)2016-01-20
少儿科学周刊·少年版(2015年11期)2015-12-17
