
博落回提取物对副猪嗜血杆菌诱导的猪肺泡巨噬细胞炎症反应及信号通路影响的研究
2022-11-05 03:06张华琦桂干北
中国预防兽医学报 2022年8期
曾 泽,张华琦,桂干北
(铜仁职业技术学院国家民委中兽药重点开放实验室,贵州 铜仁 554300)
副猪嗜血杆菌(Glaesserella parasuis,G. parasuis)属于巴氏杆菌科的嗜血杆菌属,是一种存在于猪上呼吸道的条件致病菌[1]。副猪嗜血杆菌可引起宿主以纤维性多发性浆膜炎、脑膜炎、关节炎、急性肺炎和败血症为特征的格拉瑟病(Glässer′s disease)[2]。该菌感染会导致猪的发病率和死亡率显著增加,给全球养猪业造成了重大经济损失[3]。到目前为止,可将副猪嗜血杆菌分为1~15 个血清型,但在某些国家,多达25%的分离株尚未分型[4]。在国内,最常见的副猪嗜血杆菌血清型为4 型和5 型,其次为12 型、13型和14 型,其中5 型为强毒力菌株,4 型为中等毒力菌株[5]。由于副猪嗜血杆菌血清型众多且地方性特征明显,不同的血清型菌株之间缺乏有效的交叉保护力,所以对该菌所致疫病的防治方面还存在很大困难。研究结果显示,副猪嗜血杆菌可以通过激活NF-κB 和丝裂原活化蛋白激酶(MAPK)信号通路释放炎症细胞因子,炎症细胞因子再通过旁分泌或体循环作用于组织细胞,引起炎症反应和组织损伤[6-7]。
博落回是罂粟科博落回属植物,含有多种活性生物碱,如血根碱、白屈菜红碱、原阿片碱等[8]。博落回是我国广泛使用的传统中草药,具有消炎、杀菌、杀虫、抗病毒和抗肿瘤等作用,且不易产生耐药性,是较为理想的抗生素替代品[9-10]。研究结果显示,博落回提取物可作为猪、牛、家禽和鱼等的饲料添加剂,不但能预防和治疗畜禽呼吸系统和消化系统疾病,还能促进畜禽的生长[11-12]。博落回的有效成分血根碱具有抗炎作用,能够抑制炎症反应的关键调控因子核转录因子(NF-κB)的激活[13]。从以上结果推测,博落回提取物可能会抑制副猪嗜血杆菌诱导的宿主炎症反应。基于此,本研究将不同浓度博落回提取物对猪肺泡巨噬细胞(PAMs)预处理,再将副猪嗜血杆菌感染PAMs 后,分别利用荧光定量PCR(qRT-PCR)和western blot 检测PAMs 中炎性细胞因子mRNA 的转录水平,以及NF-κB 和MAPK 信号通路相关蛋白的表达量,以确定博落回提取物对副猪嗜血杆菌诱导PAMs 中炎性细胞因子产生,以及NF-κB 和MAPK 信号通路的影响,以期为副猪嗜血杆菌病的防治提供一种新的治疗策略。
1 材料与方法
1.1 菌株、细胞和药物副猪嗜血杆菌SW124株购自中国兽医微生物菌种保藏管理中心;PAM 购自ATCC公司;博落回提取物购自湖南美可达生物资源有限公司,其中血根碱含量40%,白屈菜红碱含量20%。
1.2 主要试剂胰蛋白胨大豆琼脂(TSA)、胰蛋白胨大豆琼脂(TSB)购自北京奥博星生物技术有限公司;烟酰胺腺嘌呤二核苷酸(NAD)购自Aladdin 公司;新生牛血清、RPMI1640 培养基和胎牛血清购自Gibco 公司;PrimeScriptTMRT reagent kit、SYBR premix ExTaqTM、TaqDNA聚合酶、RNAiso plus 购自宝生物工程(大连)有限公司;PMSF、RIPA 裂解液、BCA 蛋白浓度测定试剂盒购自上海碧云天生物技术有限公司;鼠源p65 单克隆抗体(MAb)、兔源p-p65 MAb、鼠源IκBα MAb、鼠源p-IκBα MAb、兔源JNK 多克隆抗体(pAb)、兔源p-JNK pAb、兔源ERK pAb、兔源p-ERK pAb、兔源p38 pAb、兔源p-p38 pAb和兔源β-actin MAb购自美国Cell Signaling Technology公司;HRP标记的羊抗兔和羊抗鼠IgG(IgG-HRP)购自美国Abbkine公司。
1.3 实验设计将PAM 以1×107个/孔铺于24 孔板中,在37 ℃5%CO2培养24 h 后,实验组以终浓度分别为12.5 ng/mL、25 ng/mL、50 ng/mL、100 ng/mL 和200 ng/mL 的博落回提取物分别预处理PAM 4 h 后,以1×107cfu/mL 副猪嗜血杆菌感染上述各组细胞,5% CO237 ℃培养12 h。设无处理无感染的空白对照组,设TSB 培养液同样预处理PAMs 4 h 再感染副猪嗜血杆菌作为感染对照组。
1.4 各组细胞中炎性细胞因子基因转录水平的qRT-PCR 检测上述各组PAM 弃去上清液,加入RNAiso plus 收集PAM,提取细胞总RNA,反转录为cDNA作为模板,参考文献[14]中的炎性细胞因子引物,通过qRT-PCR 检测各组细胞IL-1α、IL-1β、IL-6、IL-8和TNF-α mRNA的转录水平。反应条件为:95 ℃5 min;95 ℃30 s,57 ℃31 s;共40 个循环,同时用无菌水代替cDNA 作为阴性对照,每个样品设置3 个重复,采用2-ΔΔCT法计算炎性细胞因子的相对转录水平,ΔΔCT=实验组(目的基因Ct-内参基因Ct)-对照组(目的基因Ct-内参基因Ct)。研究显示,核糖体蛋白L4(RPL4)基因能够在PAM中能够稳定转录[15],所以选用RPL4作为qRT-PCR检测的内参基因。
1.5 各组细胞中NF-κB 和MAPK 信号通路蛋白表达的western blot 检测将1.3 中的各组PAMs 弃去上清液,加入RIPA 裂解液裂解细胞,离心取上清即为细胞总蛋白,利用BCA 试剂盒测定所提取蛋白质浓度。取40 μg 蛋白质分别以p65、p-p65、IκBα、p-IκBα、JNK、p-JNK、ERK、p-ERK、p38、p-p38、β-actin 相应的MAb 和pAb 为一抗(1∶1 000),以羊抗兔或羊抗鼠IgG-HRP(1∶5 000)为二抗,经western blot 检测各信号通路相关蛋白的表达,利用Image J软件计算NF-κB 和MAPK 信号通路各蛋白条带的灰度值,采用Graphpad prism 6.0 软件制做柱状图。
1.6 数据分析本研究所有数据均利用SPSS 16.0 软件对检测数据进行单因素方差分析,采用Duncan 法进行多重比较,P<0.05 为差异有统计学意义。
2 结 果
2.1 博落回提取物对副猪嗜血杆菌诱导PAM 炎性细胞因子mRNA 转录水平影响的检测结果采用不同浓度的博落回提取物预处理PAM 4 h 后,再利用副猪嗜血杆菌感染12 h后,通过qRT-PCR检测各炎性细胞因子mRNA 转录水平。结果显示,与空白对照组相比,感染对照组细胞中炎性因子IL-1α、IL-1β、IL-6、IL-8 和TNF-α mRNA 的转录水平均显著升高(P<0.05)。与感染对照组相比,经12.5 ng/mL、25 ng/mL、50 ng/mL、100 ng/mL 和200 ng/mL 博落回提取物预处理后再感染副猪嗜血杆菌的各实验组炎性细胞因子IL-1α、IL-1β、IL-6、IL-8 和TNF-α mRNA 的转录水平均显著降低(P<0.05),且随着博落回提取物浓度的升高,上述各炎性细胞因子mRNA 转录水平降低的越多(图1)。表明博落回提取物能够抑制副猪嗜血杆菌诱导的PAM中炎性细胞因子mRNA的转录,且该抑制作用与博落回提取物的浓度呈正相关。
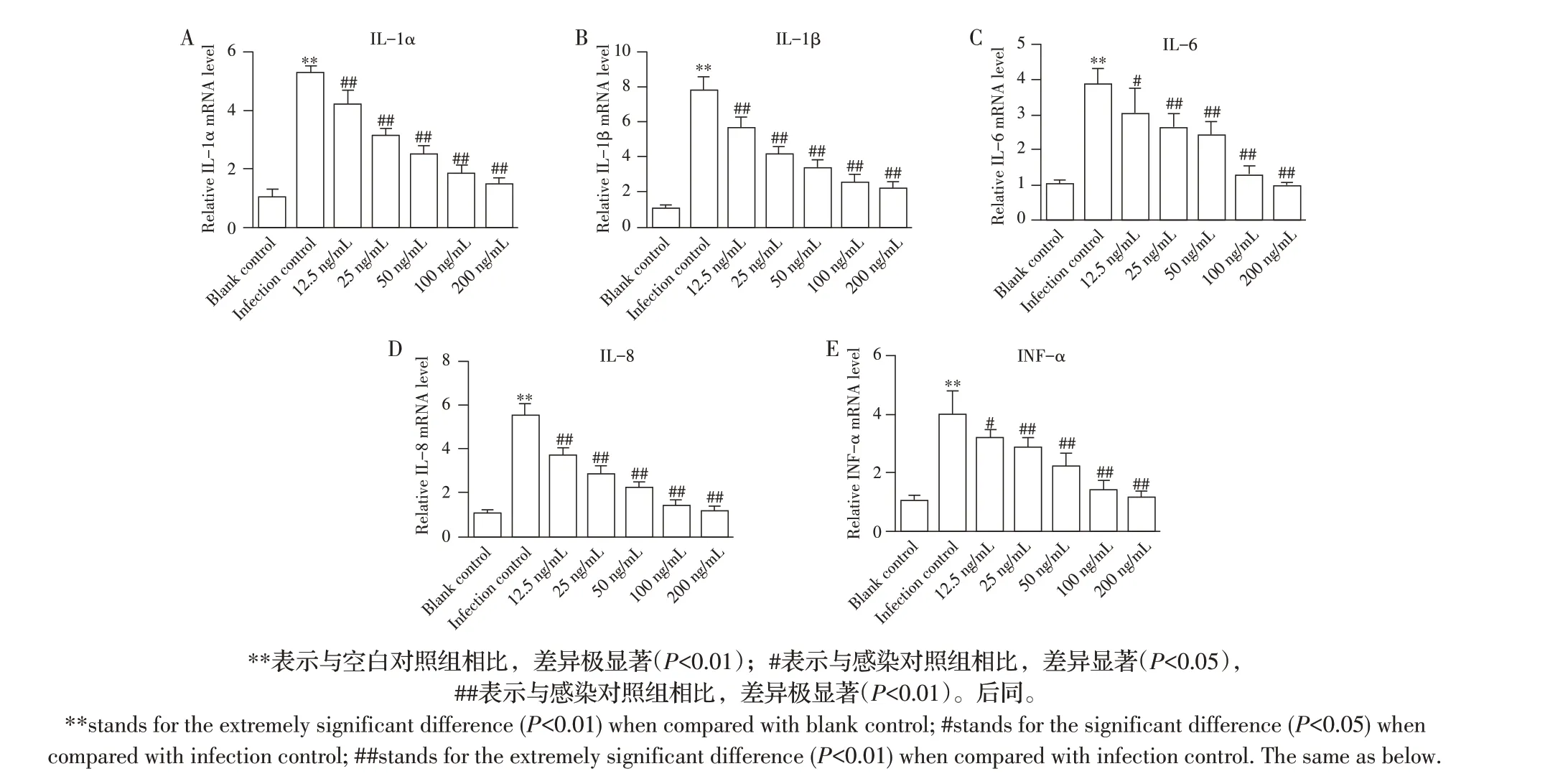
图1 博落回提取物对副猪嗜血杆菌诱导PAM后炎性因子mRNA转录水平影响的检测结果Fig.1 The effect of M.cordata extract on the mRNA transcription of inflammatory cytokines in PAM induced by G.parasuis
2.2 博落回提取物对副猪嗜血杆菌诱导PAM 中NF-κB 信号通路蛋白表达影响的检测结果采用各浓度博落回提取物预处理PAM 4 h 再经副猪嗜血杆菌感染12 h 后,通过western blot 检测NF-κB 信号通路中各蛋白的表达量。结果显示,与空白对照组相比,感染对照组p-p65和p-IκBα蛋白的表达量显著升高(P<0.05)。与感染对照组相比,经不同浓度博落回提取物预处理再经副猪嗜血杆菌感染后的PAM 中p-p65 和p-IκBα 蛋白表达量均显著降低(P<0.05),且随着博落回提取物浓度的升高二者的表达量均下降;但各组间p65 和IκBα 蛋白的表达量均无显著变化(P>0.05)(图2)。表明博落回提取物能够抑制副猪嗜血杆菌诱导PAM 中NF-κB 信号通路p-p65 和p-IκBα 蛋白的表达,且该抑制作用与博落回提取物的浓度呈正相关。
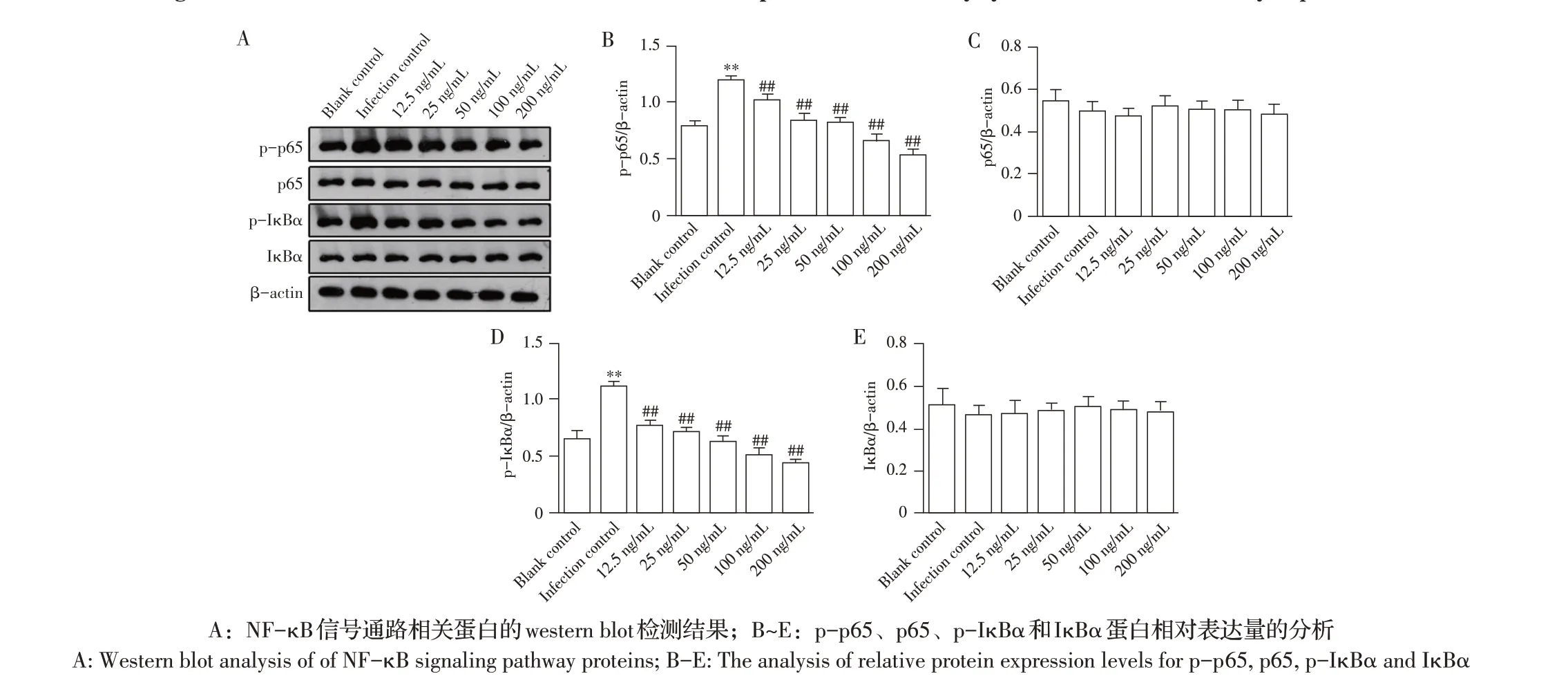
图2 博落回提取物对副猪嗜血杆菌诱导PAM后NF-κB信号通路相关蛋白表达影响的检测结果Fig.2 The effect of M.cordata extract on the protein expression of NF-κB signaling pathway in PAM induced by G.parasuis
2.3 博落回提取物对副猪嗜血杆菌诱导PAM中MAPK信号通路相关蛋白表达水平影响的检测结果将各浓度的博落回提取物预处理PAM 4 h,再经副猪嗜血杆菌感染12 h 后,通过western blot 检测MAPK信号通路各蛋白的表达水平。结果显示,与空白对照组相比,感染对照组中p-JNK、p-ERK 和p-p38 的蛋白表达水平均显著升高(P<0.05)。与感染对照组相比,经不同浓度博落回提取物预处理再感染副猪嗜血杆菌后的各实验组中p-JNK 和p-p38 的蛋白表达水平均显著降低(P<0.05);除了12.5 ng/mL 博落回提取物处理的PAM 外(P>0.05),其余浓度博落回提取物预处理再感染副猪嗜血杆菌后的PAM 中p-ERK 的蛋白表达水平均显著降低(P<0.05)。各组PAM 中JNK、ERK 和p38 的蛋白表达水平均无显著性变化(P>0.05)(图3)。表明博落回提取物能够抑制副猪嗜血杆菌诱导PAM 中MAPK 信号通路p-JNK、p-ERK 和p-p38 蛋白的表达,且该抑制作用基本与博落回提取物的浓度呈正相关。
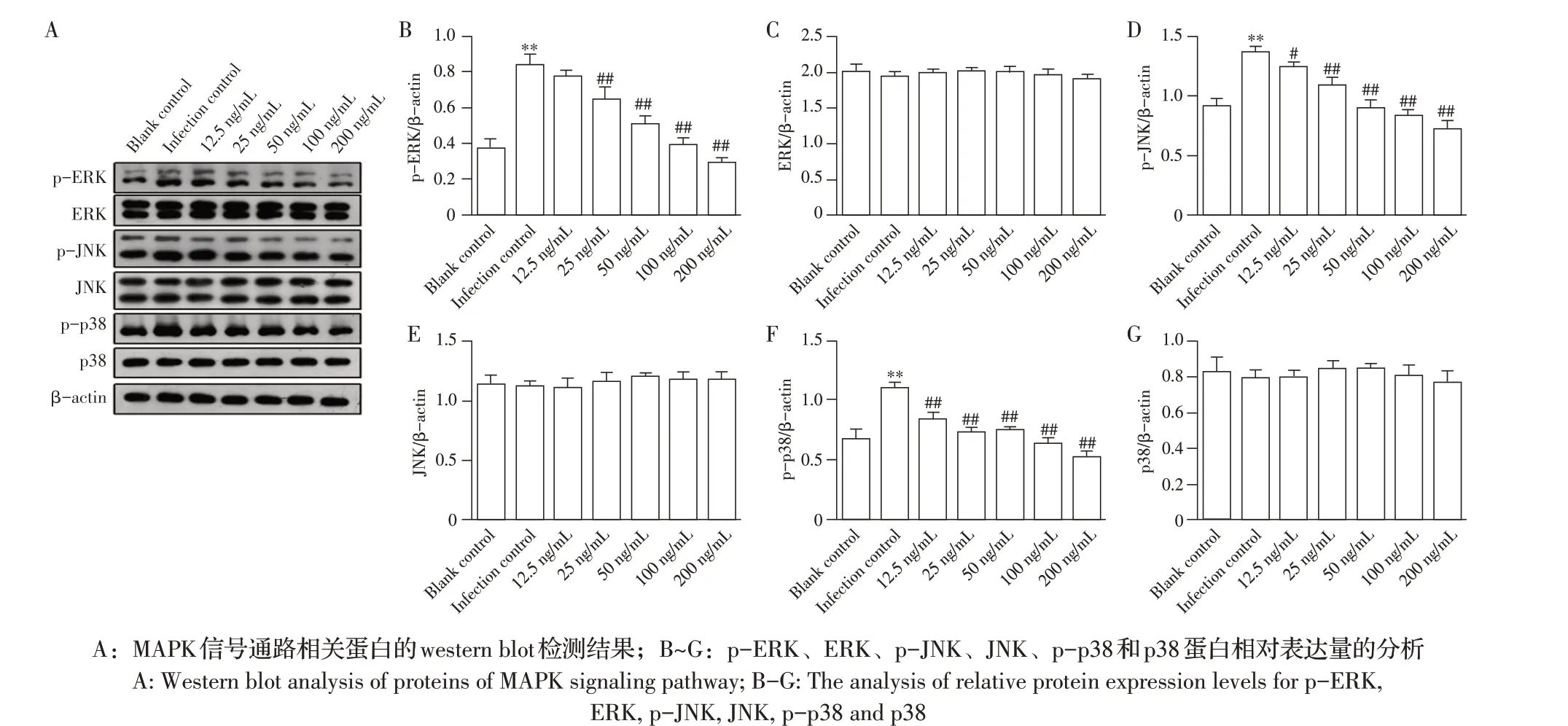
图3 博落回提取物对副猪嗜血杆菌诱导PAM后MAPK信号通路相关蛋白表达影响的检测结果Fig.3 The effect of M.cordata extract on the protein expression of MAPK signaling pathway in PAM induced by G.parasuis
3 讨 论
副猪嗜血杆菌的感染会引起宿主的炎症反应,主要以呼吸道症状、胸腔和腹腔大量纤维素性物质渗出、心包积液和关节炎为特征性症状,发病率和死亡率较高,给养猪业造成重大的经济损失[16]。研究表明副猪嗜血杆菌可以激发宿主先天免疫反应并诱导猪肾上皮细胞(PK-15)、猪脑微血管内皮细胞(PBMEC)、仔猪单核吞噬细胞(PMNP)、猪主动脉血管内皮细胞(PAVECs)和PAM 产生炎性细胞因子[17-20]。TNF-α、IL-1β 和IL-6 等炎性细胞因子是炎症反应的重要标志物,且这些致炎因子会引起宿主组织的损伤[10,21]。因此,抑制宿主的炎症反应可能是预防和治疗副猪嗜血杆菌病的有效方法。博落回具有抗菌、抗炎和抗氧化等多种药理作用,已经在畜禽生产中得到较为广泛的应用[11-12]。为了研究博落回提取物对副猪嗜血杆菌感染引起的炎症反应的影响,本研究分析了副猪嗜血杆菌感染博落回提取物预处理后的PAM 中IL-1α、IL-1β、IL-6、IL-8 和TNF-α 的mRNA 转录水平,结果显示副猪嗜血杆菌刺激PAM 后上述细胞因子的mRNA 转录水平均显著升高,但经博落回提取物预处理后的PAM 中上述细胞因子的mRNA 转录水平均受到显著抑制,说明博落回提取物可以抑制副猪嗜血杆菌刺激引起的宿主细胞炎症反应。
NF-κB 和MAPK 信号通路能够调节宿主炎性细胞因子的表达,参与炎症反应和免疫调节等多种生物学反应[7,22-23]。NF-κB 转录因子家族由5 个亚基组成:p65、RelB、c-Rel、NF-κB1 和NF-κB2,其中NF-κB 主要参与细胞对外界刺激的响应[24]。MAPK 家族蛋白包括p38、JNK 和ERK,MAPK 是NF-κB 信号通路的主要介质,因此MAPK 在炎症反应的调节中至关重要[25-26]。已有研究证实NF-κB 和MAPK 信号通路参与副猪嗜血杆菌感染诱导的宿主炎症反应的激活[6-7]。所以本研究检测了副猪嗜血杆菌感染博落回提取物预处理后的PAM 中NF-κB 信号通路蛋白p65、p-p65、IκBα 和p-IκBα 及MAPK 信号通路蛋白p38、p-p38、ERK、p-ERK、JNK和p-JNK的表达水平。结果显示,副猪嗜血杆菌刺激PAM 后p-p65、p-IκBα、p-p38、p-ERK 和p-JNK 蛋白的表达水平均显著升高,但博落回提取物能够抑制副猪嗜血杆菌刺激PAM后上述蛋白的表达,表明博落回提取物可以通过抑制NF-κB 和MAPK 信号通路从而抑制副猪嗜血杆菌诱导的炎症反应。后续将对博落回提取物对副猪嗜血杆菌在猪体内感染中的抗炎作用及作用机制作深入研究。
本研究首次证实了博落回提取物能够通过抑制副猪嗜血杆菌刺激PAM 后NF-κB 和MAPK 信号通路的激活从而抑制副猪嗜血杆菌诱发的炎症反应。表明博落回提取物可以抑制副猪嗜血杆菌感染所致的炎症反应,本研究为防治副猪嗜血杆菌病提供实验依据。
猜你喜欢
中国现代医生(2022年21期)2022-08-22
材料与冶金学报(2022年2期)2022-08-10
纺织标准与质量(2022年3期)2022-08-10
健康体检与管理(2022年4期)2022-05-13
检验医学与临床(2022年9期)2022-05-13
新疆医科大学学报(2022年1期)2022-01-27
建材发展导向(2021年23期)2021-03-08
诗潮(2020年4期)2020-04-26
东坡赤壁诗词(2018年6期)2018-12-22
当代体育·扣篮(2014年8期)2014-05-23
