
马铃薯块茎蛾取食胁迫下马铃薯根际土壤微生物代谢功能多样性变化
2023-12-11 10:02唐芬芬杨伟克张祖芸
河南农业科学 2023年11期
唐芬芬,杨伟克,谢 昆,张祖芸,李 娜
(1.红河学院 生物科学与农学学院,云南 蒙自 661199;2.云南省农业科学院 蚕桑蜜蜂研究所,云南 蒙自 661199)
根际土壤微生物种类众多,是最复杂的生态系统之一,因此也被誉为植物的第二套基因组[1],它们在植物的营养吸收[2-4]、生长发育[5]、免疫[6]及抗病性[7-9]等方面具有重要作用。过去几十年,植物的防御机制研究主要集中在相关抗性基因方面[10-11],关于植物遗传对许多代谢防御途径的影响已经很清楚,例如,调节次生代谢产物产生的植物途径极大地影响了田间环境中草食动物对植物的损害,是极其重要的防御机制之一[12-13]。近几年研究发现,植物根际土壤微生物还能影响植物对植食昆虫的抗性[14-17],进一步揭示了植物-微生物-植食昆虫三者之间的关系。
马铃薯块茎蛾(Phthorimaea operculellaZeller)是马铃薯的重要害虫之一,不仅取食马铃薯植株,还能危害块茎[18-19]。此外,该虫对烟草、辣椒等茄科作物也造成危害[20-21]。化学农药是防控该虫害的主要方式,但化学农药的广泛使用使该虫对拟除虫菊酯类及有机磷类等杀虫剂都产生了不同程度的抗性[22]。目前,国内外对马铃薯块茎蛾的防控研究主要集中在综合治理方面,如栽培管理[23]、间套作[24]、化学信息素[25]、生物杀虫剂[26-28]、植物抗性诱导剂[29]、生物防治[30]等,这些方法虽取得了一定成效,但仍需开展防治马铃薯块茎蛾的其他途径研究。目前,关于马铃薯根际微生物是否也与植株抗虫性相关以及马铃薯块茎蛾取食胁迫对马铃薯根际微生物群落的影响研究尚未见报道。因此,采用Biolog 技术分析马铃薯块茎蛾取食胁迫下云南省2个主栽马铃薯品种根际微生物代谢功能的变化,明确马铃薯块茎蛾取食胁迫对其根际微生物的影响,为深入探究马铃薯块茎蛾、马铃薯以及根际微生物之间的相互作用机制提供理论基础,并为马铃薯抗虫性机制研究以及马铃薯病虫害防控提供理论依据和新思路。
1 材料和方法
1.1 试验材料
Biolog ECO 板购自美国BIOLOG 公司;马铃薯品种为合作88、丽薯6 号;供试土壤采自红河学院种植基地,一次性采集并充分混匀、去除杂质,过孔径0.6 mm筛后备用。
1.2 马铃薯种植
将上述过筛的土壤分装于洗净的花盆里,每盆种植马铃薯块茎一个(保留一个芽眼)。然后置于温度为(26±1)℃、湿度为(75±5)%、光周期L/D=16 h/8 h 的光照培养箱内培养(恒立国际有限公司,型号RG-300),在此期间无任何虫源危害,不施加任何化肥、农药,按需进行常规浇水。采集土壤样品前3 d不浇水。
1.3 马铃薯块茎蛾胁迫处理
收集马铃薯块茎蛾同一天卵块,待其孵化后,接于出苗45 d 的马铃薯植株上,分别于马铃薯块茎蛾取食3、7、14 d 时采集土壤,同时以无害虫取食胁迫的马铃薯植株为对照组。采集方法为抖落法。马铃薯植株取出后,将与根系结合不紧密的部分土壤抖落,然后用无菌毛笔将与根结合紧密的土壤扫落于无菌牛皮纸上。分别将2个品种取食胁迫各时段(3、7、14 d)的土壤每3 盆混在一起作为一个样品,每个处理3 个重复,即2 个胁迫处理54 盆,对照54 盆,共108 盆。收集样品用于微生物代谢功能多样性相关数据的测定。
1.4 根系微生物代谢功能测定
根际微生物变化与根系分泌物利用的关系可以在一定程度上通过Biolog ECO 微孔板分析来反映,因此,本研究中对土壤微生物代谢功能多样性分析采用Biolog 生态板的方法进行测定。采用的Biolog ECO板有31种单一碳源,分为六大类:碳水化合物类、氨基酸类、羧酸类、多聚物类、酚酸类、胺类。
测定方法参照左强等[31]的报道,略有改进。不同处理分别称取混合土壤5.0 g,加入90 mL 无菌的0.85% NaCl 溶液,放置于25 ℃摇床200 r/min 振荡30 min 后于4 ℃冰箱静置10 min,用移液枪吸取上清液200 μL 稀释1 000 倍,然后取150 μL 稀释液添加至Biolog ECO 板的微孔中,最后将Biolog ECO 板培养于25 ℃恒温培养箱中,分别在培养4、12、24、48、72、96、120、144、168、192 h 时取出Biolog ECO板,在Biolog EmaxTM自动读盘机上读取590 nm 波长的光密度值,选取测定趋于平稳的96 h 光密度值进行数据分析。
1.5 数据处理与分析
土壤微生物可与Biolog ECO 板微孔内的染料四唑反应,产生颜色变化,通过颜色深浅可判断微生物对碳源的利用能力,通过平均颜色变化率(Average color change rate,AWCD)可反映土壤微生物的代谢活性情况,即AWCD 值越大,土壤微生物的代谢活性越强。其计算公式为AWCD=∑(Ci-Ri)/n,其中:Ci为每个孔的光密度值;Ri为对照孔的光密度值;n=31,为Biolog ECO板的孔数。
微生物多样性指数利用Excel 计算。评估物种的丰富度用Shannon指数(H),计算公式为H=∑Pi/lnPi,Pi为第i孔的相对吸光值与整个微平板所有孔中相对吸光值总和的比值;Simpson 多样性指数(D)用于评估优势度,计算公式为D=1-∑Pi2[32]。
数据分析采用Excel 2010 和SPSS 20.0 软件进行,不同处理间在0.05 水平下的差异显著性通过邓肯氏多重比较法检验。
2 结果与分析
2.1 马铃薯块茎蛾取食胁迫对马铃薯根际微生物代谢活性的影响
根系微生物代谢活性可用AWCD 值衡量,结果(图1)显示,经马铃薯块茎蛾幼虫取食后,马铃薯品种合作88、丽薯6 号的AWCD 值均高于对照,两品种的AWCD 值均在取食胁迫7 d 达到最高,且显著高于对照(P<0.05)。马铃薯块茎蛾幼虫取食胁迫3、7、14 d,合作88 AWCD 值比对照分别提高了3.9%、32.8%、11.8%;而丽薯6 号则比对照分别提高了23.1%、10.1%、35.1%。

图1 马铃薯块茎蛾取食后马铃薯根际土壤微生物的代谢活性变化Fig.1 Variation of metabolic activity of potato rhizosphere soil microorganisms after feeding by P.operculella
2.2 马铃薯块茎蛾取食胁迫对马铃薯根际微生物多样性的影响
微生物多样性分析结果(表1、表2)表明,在马铃薯块茎蛾幼虫取食胁迫处理第7 天,两品种的Shannon 指数和Simpson 指数均最高,即根际微生物多样指数均达到最高,但丽薯6 号胁迫处理此时的2 个指数与对照相比均未达到显著水平(P<0.05)。合作88 和丽薯6 号根际微生物的Shannon 指数在取食胁迫第3、7、14 天相对于对照分别提高了1.9%、2.5%、1.2%和2.8%、0.6%、3.1%,Simpson 指数则分别对应提高了6.4%、15.0%、6.8%和12.7%、4.4%、18.5%。

表1 马铃薯块茎蛾取食对马铃薯根际土壤微生物群落Shannon多样性指数的影响Tab.1 Effects of feeding by P.operculella on Shannon diversity index of soil microbial community in potato rhizosphere

表2 马铃薯块茎蛾取食对马铃薯根际土壤微生物群落Simpson多样性指数的影响Tab.2 Effects of feeding by P.operculella on Simpson diversity index of soil microbial community in potato rhizosphere
2.3 马铃薯块茎蛾胁迫下根际微生物对碳源利用的差异
通过测定微生物对Biolog ECO 板上各类碳源的平均吸光度值,评价微生物对不同碳源的利用能力,分析了马铃薯块茎蛾幼虫取食胁迫不同时间下根际微生物对31 种碳源即归类为六大类碳源利用的差异。从图2 可以看出,无论是合作88 还是丽薯6号在取食胁迫第7天,其根际微生物对6类碳源的利用均达到最大。合作88 在胁迫处理第3 天,根际微生物相对于对照仅对氨基酸的利用达到显著提高水平(P<0.05);在胁迫处理第7 天,与对照相比,合作88 的根际微生物提高了对6 类碳源的利用,分别为碳水化合物类、氨基酸类、羧酸类、多聚物类、酚酸类和胺类,其中对碳水化合物类和羧酸类化合物的利用能力与对照相比达到显著水平(P<0.05);在胁迫处理14 d,合作88 根际微生物与对照相比显著提高了对氨基酸类、多聚物类和胺类的利用(P<0.05)(图2a)。
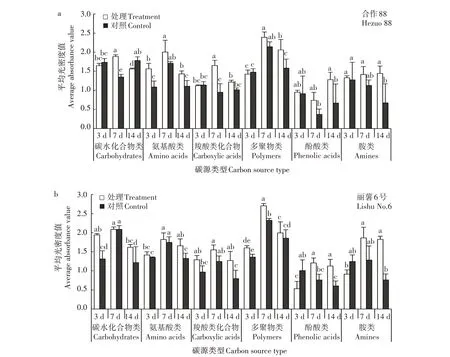
图2 马铃薯块茎蛾取食后马铃薯根际土壤微生物在各时段对6类碳源的利用情况Fig.2 Utilization of six carbon sources by microorganisms of potato rhizosphere soil at different periods after feeding by P.operculella
而丽薯6 号在处理第3 天,与对照相比其根际微生物提高了对4 类碳类的利用,分别为碳水化合物类、氨基酸类、羧酸类、多聚物类,但对酚酸类和胺类化合物的利用则低于对照,其中氨基酸类、羧酸类和多聚物类未达到显著水平(P<0.05);在处理第7 天时,丽薯6 号的根际微生物对多聚物类和酚酸类化合物的利用情况与对照相比达到显著水平(P<0.05);在胁迫处理14 d,丽薯6 号根际微生物对6 类碳源的利用情况与对照相比,仅有多聚物类未达到显著水平(P<0.05)(图2b)。
综上所述,取食胁迫处理后,马铃薯根际微生物对碳源的利用随时间推移呈现动态变化,2 个品种的根际微生物对碳源的利用不同。总体上显示,马铃薯块茎蛾取食胁迫处理提高了马铃薯根际微生物对各类碳源的利用,其中取食胁迫第7 天时马铃薯根际微生物对碳源的利用达到最高。
2.4 马铃薯根际微生物对碳源利用的主成分分析
为了进一步了解马铃薯块茎蛾取食下马铃薯不同品种根际土壤微生物群落功能的差异,对不同处理下培养96 h 的根际土壤微生物进行31 种碳源利用的主成分分析,按照载荷值大于0.18 或者小于-0.18 的标准[33],进行重要碳源的筛选。从表3 看出,影响品种合作88 的重要碳源,主成分1 上有11种,分别是丙酮酸甲脂、吐温40、肝糖、I-赤藻糖醇、D,L-α-甘油、衣康酸、γ-羟基丁酸、腐胺、L-丝氨酸、L-苏氨酸、L-苯基丙氨酸;主成分2 上有10 种,分别为D-纤维二糖、β-甲基D-葡萄糖苷、葡萄糖-1-磷酸盐、D-半乳糖内酯、衣康酸、α-丁酮酸、D-苹果酸、L-精氨酸、苯乙基胺、L-丝氨酸。
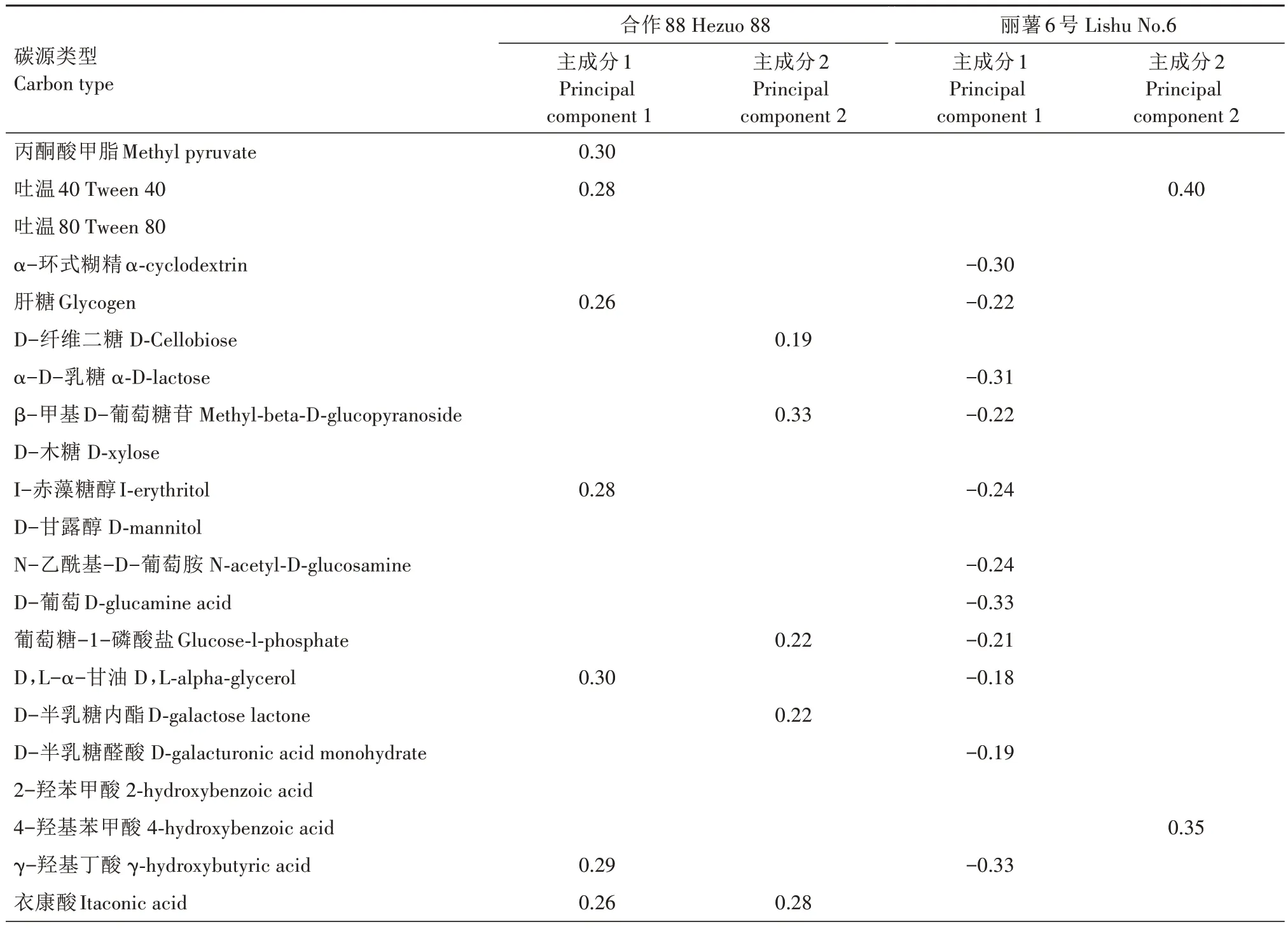
表3 31种碳源在主成分1(PC1)、主成分2(PC2)上的载荷(Loading>0.18和Loading<-0.18)Tab.3 Loads of 31 carbon sources on first two principal components(PC)(the loading is>0.18 or <-0.18)
影响丽薯6 号的重要碳源在主成分1 上有12种,分别为α-环式糊精、肝糖、α-D-乳糖、β-甲基D-葡萄糖苷、I-赤藻糖醇、N-乙酰基-D-葡萄胺、D-葡萄胺酸、葡萄糖-1-磷酸盐、D-半乳糖醛酸、4-羟基苯甲酸、甘氨酰-L-谷氨酸、L-丝氨酸;主成分2上有7 种,分别为吐温40、4-羟基苯甲酸、α-丁酮酸、L-精氨酸、腐胺、苯乙基胺、甘氨酰-L-谷氨酸。
从排序图(图3)来看,合作88 主成分1 变异为28.3%,主成分2 变异为13.6%,总变异为41.9%,且各时间段处理组和对照组均分布于不同象限;丽薯6 号主成分1 的变异为37.5%,主成分2 的变异为8.5%,总变异为46.0%,各时间段处理组和对照组也均分布于不同象限。可见,取食胁迫改变了根际微生物对碳源的利用。
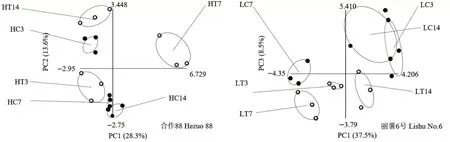
图3 马铃薯块茎蛾取食后马铃薯根际土壤微生物碳源利用的主成分变异分析Fig.3 Principal component variation analysis of microbial carbon source utilization in potato rhizosphere soil after feeding by P.operculella
3 结论与讨论
本研究利用Biolog 技术分析研究了丽薯6 号、合作88 两个马铃薯品种在马铃薯块茎蛾胁迫下根系土壤微生物群落多样性及对土壤碳源利用的情况。土壤碳源利用AWCD 值可衡量微生物对相应碳源的利用能力。经马铃薯块茎蛾取食3、7、14 d的2 个马铃薯品种根际土壤样品,在培养96 h 时根际微生物对碳源的利用能力均高于对照,且随取食后时间延后出现先上升后下降的趋势,代谢速率与微生物的增长规律吻合,即慢-快-慢的代谢速率[34],且均在取食胁迫第7 天微生物对碳源的利用达到最大。结果说明,在马铃薯块茎蛾取食胁迫下,植株根际土壤微生物对碳源的代谢活性和利用能力增强。
微生物群落功能多样性变化可以通过微生物多样性指数来反映,Shannon 和Simpson 指数是最常用的指标之一,值越高说明群落多样性越高,物种越丰富[35]。对马铃薯块茎蛾取食胁迫下2 个马铃薯品种根际微生物多样性的分析结果表明,取食胁迫处理的Shannon和Simpson 指数值均高于对照,说明在马铃薯块茎蛾取食胁迫下,2 个马铃薯品种根际微生物的生物多样性得到提高。
本研究分析了马铃薯根际微生物对Biolog Eco板31种碳源,即归属于碳水化合物类、氨基酸类、羧酸类、多聚物类、酚酸类、胺类6类碳源的利用情况。与对照相比,马铃薯块茎蛾取食胁迫明显提高了2个品种根际土壤微生物对6类碳源的利用,且趋势基本一致,说明取食胁迫后微生物的群落更为丰富和多样,与上述结果一致。糖类、氨基酸类和酚酸类化合物是植物根系分泌物的主要成分[36],在根际与周围微生物的相互作用中发挥着重要作用。研究表明,在根系分泌物的作用下,具有趋化性的细菌或真菌能够在根际大量聚集和繁殖[37]。对马铃薯根际微生物的碳源利用情况进行主成分及聚类分析,结果表明,马铃薯块茎蛾取食胁迫处理与对照相比,改变了根际微生物对碳源的利用,说明微生物群落结构和生物多样性发生了变化。同时研究还发现,未受害虫取食胁迫的对照组2 个马铃薯品种的根际微生物对碳源利用的主成分不同,说明群落有差别,这与已有研究报道认为植物种类对根际微生物群落有影响的结论相符,即使是相同物种,基因型不同也会导致微生物群落的差异[38]。
植物、微生物与植食昆虫之间存在着密切关系,植物根系能感知微生物的存在,根系分泌物则是植物根系与土壤之间进行信息交流和能量传递的主要媒介[39]。研究表明,昆虫取食植物能改变植物根系分泌物的组成和数量[40]。而植物可通过根系分泌生物活性分子来改变土壤微生物区系,诱导菌群繁殖,从而影响下一代植物的生长和抗虫性[41-42],如HU 等[43]研 究 发 现,草 地 贪 夜 蛾(Spodoptera frugiperda)取食玉米叶片后会诱导玉米根际分泌属于防御性次生代谢产物的苯并恶嗪类化合物,然后通过调节土壤改变根际土壤微生物群落,最后使茉莉酸信号增强,叶片防御能力提高。另外,BADRI 等[44]研究表明,土壤微生物能通过影响植物挥发物,从而影响昆虫的取食选择性,保护植物免受昆虫的危害。本研究中,在马铃薯块茎蛾的取食胁迫下,2 个马铃薯品种植株根际土壤微生物的生长和群落结构均发生改变,总体表现为功能多样性提高,很可能是由于马铃薯块茎蛾取食诱导根系分泌物发生改变,最终造成土壤微生物群落的结构多样性改变,形成了有益于自身生长的土壤微生态环境。同时,在马铃薯取食胁迫下,2个品种的根际土壤微生物功能多样性存在一定差异,说明根际微生物群落结构和功能多样性很可能受植物品种抗虫性的影响[45]。根系微生物的这些改变是否与植株的抗虫性有关,还值得进一步的研究。此外,由于Biolog 技术限制,本研究仅发现马铃薯块茎蛾胁迫下根际土壤微生物发生了改变,但是具体哪一类微生物发生了改变,它们的改变有怎样的作用,还需深入探究。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
中国土壤与肥料(2021年5期)2021-12-02
今日农业(2021年15期)2021-11-26
昆钢科技(2021年6期)2021-03-09
天然产物研究与开发(2018年3期)2018-05-07
现代园艺(2017年19期)2018-01-19
中国蔬菜(2016年8期)2017-01-15
电源技术(2016年9期)2016-02-27
云南中医学院学报(2015年2期)2015-07-31
武夷学院学报(2014年5期)2014-07-19
