
探索学习对局灶性脑梗死大鼠梗死灶周围皮质巢蛋白及突触素表达的影响
2010-07-16 02:16槐雅萍杨永轩贾子善郭宗成贾新凤
中国全科医学 2010年5期
槐雅萍,杨永轩,贾子善,郭宗成,贾新凤
本文要点
1 探索学习大鼠脑梗死灶周围皮质巢蛋白阳性神经元数在造模后第7、14天明显多于手术对照组,提示探索学习可能通过增强脑梗死大鼠巢蛋白的表达,来保护神经元免受缺血缺氧损伤,促进脑功能的重组。
2 除大脑室管膜下区、海马齿状回外,海马 CA1~4区、缺血灶周围皮质也可见巢蛋白的表达。推测脑缺血损伤可能刺激脑内成熟的间质细胞或星形胶质细胞返回到神经干细胞状态,重新分化成神经元。
3 探索学习与突出素的表达呈正相关;大鼠经过探索学习训练,随着时间的延长,突出素的反应产物明显增多。
中枢神经系统可塑性变化是康复治疗的机制之一。半个世纪前,就有学者提出神经皮质的连接可因个体的经历 (如学习、感觉刺激、物理刺激等)而发生改变,脑功能代表区的改变也是如此。探索学习是指动物主动去接受新的信息而改变自身行为、适应新环境的过程,是不断用新记忆取代旧记忆的过程[1],主要用来训练动物的学习、记忆、空间辨别、觅食等本能性行为能力,促进受损的行为学功能的恢复。有研究表明探索学习能明显促进脑梗死大鼠的行为学恢复[2],但对脑卒中后行为学能力恢复的分子作用机制尚不清楚。本研究拟观察探索学习对局灶性脑梗死大鼠梗死灶周围皮质巢蛋白 (nestin)及突触素 (synaptophysin,SYP)表达的影响,探讨探索学习对脑梗死后功能恢复影响的机制。
1 材料与方法
1.1 实验动物 清洁级雄性 SD大鼠 70只,3~4月龄,体质量 230~260 g,购自华中科技大学同济医学院实验动物学部。合格证号 SCXK(鄂)2004—0007。将动物随机分为大脑中动脉栓塞 (MCAO)模型组60只,假手术组 10只。
1.2 试剂与仪器 戊巴比妥钠购自上海生工公司;多聚甲醛(分析纯)购自天津市化学试剂研究所;兔抗大鼠巢蛋白多克隆抗体购自北京中杉金桥;小鼠抗大鼠突触素多克隆抗体购自武汉博士德;笔式高温电灼器 (北京贝林电子有限公司);手术显微镜 (德国 Leica公司);光学显微镜 (日本 OLYMPUS公司);图像分析管理系统 (美国 Media Cybernetics公司)。
1.3 方法
1.3.1 制备 MCAO模型及分组 参照 Bederson等[3]的方法制备 MCAO模型。术后 24 h将大鼠随机分为探索学习组 30只,手术对照组 30只。假手术组 10只,不电凝大脑中动脉,其余步骤与手术组相同。
1.3.2 造模后饲养环境 (1)假手术组:饲养于标准笼,每笼 5只。(2)手术对照组:饲养于标准笼,每笼 5只。(3)探索学习组:15只一组,饲养于由一个圆笼和一个方笼组成的迷宫笼[2]。直径 500 mm的圆笼与 640 mm×480 mm×120 mm的方笼中间通过两通道相连 (圆笼:中间由丝网分隔,一侧为进食区,一侧为饮水区;方笼:由丝网分隔形成通道宽80 mm×80 mm的迷宫,迷宫由易到难,每周变换 1次)。
1.3.3 标本的制备 分别于术后第 1、7、14、28天从手术对照组和探索学习组随机取5只大鼠;假手术组于术后第7天随机取 5只大鼠,第 28天随机取 5只大鼠。用 10%水合氯醛腹腔内注射麻醉后固定于手术台,剪开胸腔,暴露心脏,用 9号针头插入左心室,灌注 0.9%氯化钠溶液,待右心耳膨起,剪开右心耳让血液流出,至右心耳流出液为无色透明时开始用4%多聚甲醛灌注固定,先快后慢,每只大鼠约灌注 300 ml,总量在 20~30 min内灌完。固定后立即剖颅取脑,在视交叉处切开,冠状切取 2 mm厚包含梗死灶区域及左右半球的组织块浸于 4%多聚甲醛中再固定 4 h。经常规梯度乙醇脱水,二甲苯透明,石蜡包埋。连续冠状切片,切片厚 5μm。
1.3.4 SP法检测巢蛋白和突触素的表达 组织切片常规脱蜡,水化。PBS冲洗 3次,每次 3 min(3×3 min,下同);0.3%过氧化氢室温孵育 10 min,消除内源性过氧化物酶;PBS冲洗 2×3 min;热修复 30 min;PBS冲洗 3×3 min;滴加溶液 A(蛋白阻断液),室温孵育 10 min,减少非特异性背景染色;PBS冲洗 3×3 min;滴加一抗 (巢蛋白稀释比例为1∶400;突触素稀释比例为 1∶500),置 20%甘油湿盒 4℃恒温过夜;PBS冲洗 3×3 min;滴加溶液 B(生物素标记的二抗),室温孵育 10 min;PBS冲洗 3×3 min;滴加溶液 C(过氧化物酶标记的链霉菌抗生素蛋白),室温孵育 10 min;PBS冲洗 3×3 min;DAB显色;自来水充分冲洗;苏木素复染;梯度乙醇脱水,二甲苯透明,中性树胶封片。PBS代替一抗做阴性对照。
1.3.5 染色结果判定 (1)巢蛋白染色结果判定:每只大鼠选取 3张切片,每张切片选取 3个 10×40倍视野,用多功能真彩色细胞图像分析管理系统计数阳性细胞数目。(2)突触素染色结果判定:每张切片在梗死灶周围皮质随机选取 3个10×40倍视野,用多功能真彩色细胞图像分析管理系统在同一光强度下测量其突触素吸光度 (optical density,OD)值,同时测量同一张切片胼胝体的 OD值作为背景 OD值,用矫正吸光度 (COD,COD=实测值 -背景值)值进行比较和分析,以避免染色过程中非特异性染色所造成的误差。
1.4 统计学方法 各组数据以 (x±s)表示,数据处理采用SPSS12.0统计分析软件,所有数据进行正态性及方差齐性检验后,组间比较采用单因素方差分析,P<0.05为差异有统计学意义。
2 结果
2.1 梗死灶周围皮质巢蛋白表达的时程变化 光学显微镜观察发现:梗死灶周围皮质巢蛋白阳性细胞密集。部分巢蛋白阳性细胞胞体较大呈椭圆形或纺锤形,胞质呈棕褐色,有少量突起,细胞核较大。假手术组皮质神经元胞质未见巢蛋白阳性表达。探索学习组与手术对照组梗死灶周围皮质巢蛋白阳性细胞数在脑梗死后第 1天即有增加,第 7天达到高峰,染色最深,第 14天和第 28天阳性细胞逐渐减少,染色变淡。脑梗死后第7天和第 14天,探索学习组大鼠梗死灶周围皮质巢蛋白阳性细胞较手术对照组明显增多,差异有统计学意义 (P<0.01),而第 28天时两组比较差异无统计学意义 (P>0.05,见表 1、图 1~3)。

表 1 各组大鼠梗死灶周围皮质巢蛋白阳性神经元数比较 (x±s,×400)Table 1 Expression of nestin in rats among three groups
2.2 梗死灶周围皮质突触素表达的时程变化 突触素免疫反应产物为棕黄色的点状或细颗粒状物,分布密集,部分呈板层样分布。有些颗粒围绕在细胞周围,衬托出细胞轮廓。假手术组皮质可见突触素呈点状或颗粒样。探索学习组与手术对照组梗死灶周围皮质突触素表达在造模后第 1天无明显变化,两组与假手术组比较差异均无统计学意义 (P>0.05),两组之间比较差异亦无统计学意义 (P>0.05)。第 7天时突触素表达增多,探索学习组与手术对照组比较差异无统计学意义 (P>0.05),而两组与假手术组比较差异均有统计学意义 (P<0.01)。第 14天时突触素表达明显增强,第 28天时最强,且颗粒较粗大,并出现深染的颗粒性产物,密集的小颗粒呈带状。探索学习组突触素表达在第 14、28天较手术对照组增强,差异有统计学意义 (P<0.01,见表 2、图 4~6)。
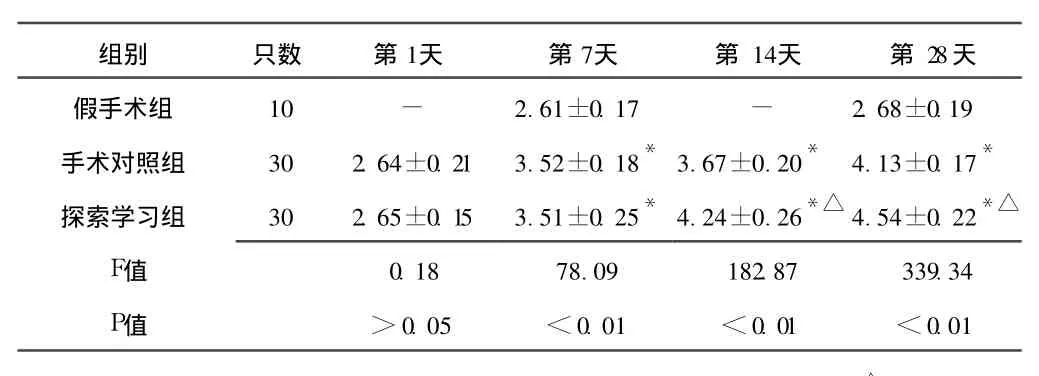
表 2 各组大鼠梗死灶周围皮质突触素光密度值比较 (x±s, ×400)Table 2 Expression of synaptophysin in rats among three groups
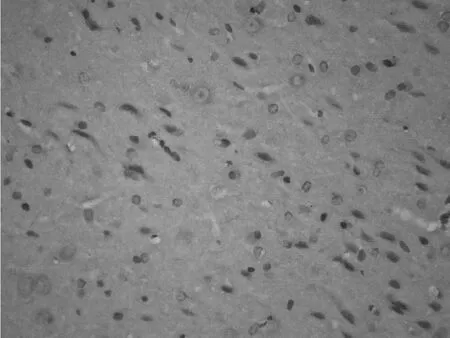
图 1 假手术组大鼠皮质巢蛋白的表达Figur e 1 Expression of nestin in cortex in sham group(IHC 400×)
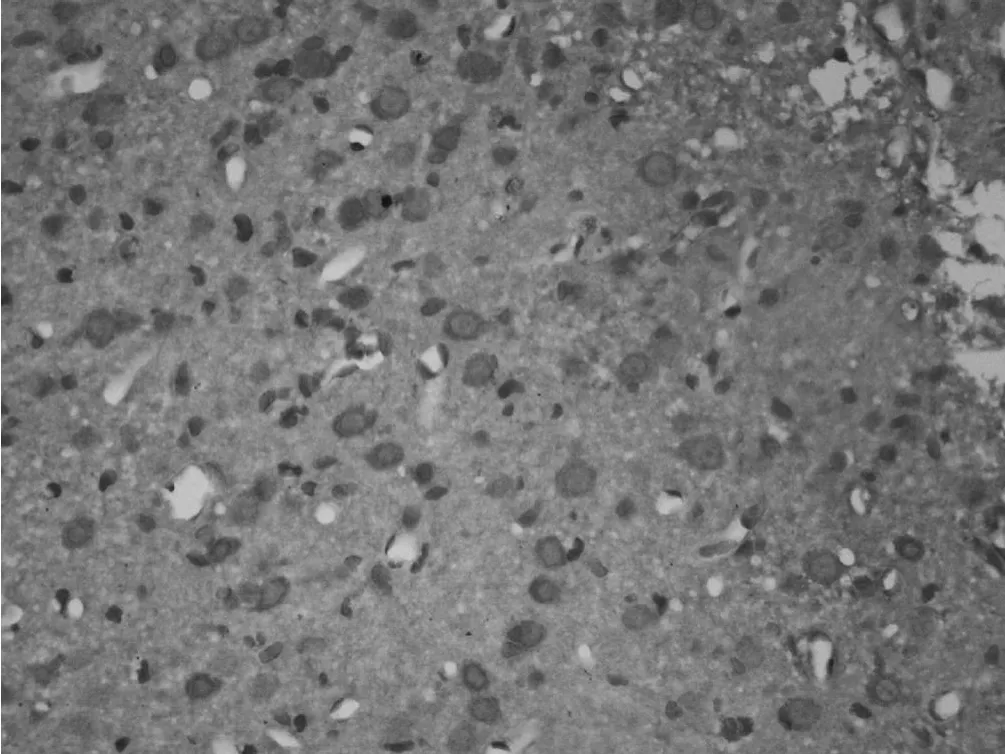
图 2 手术对照组大鼠在术后第 7天时皮质巢蛋白阳性表达情况Figur e 2 Expression of positive nestin neuron in peri-ischemic cortex in control group at 7 day after MCAO(IHC 400×)
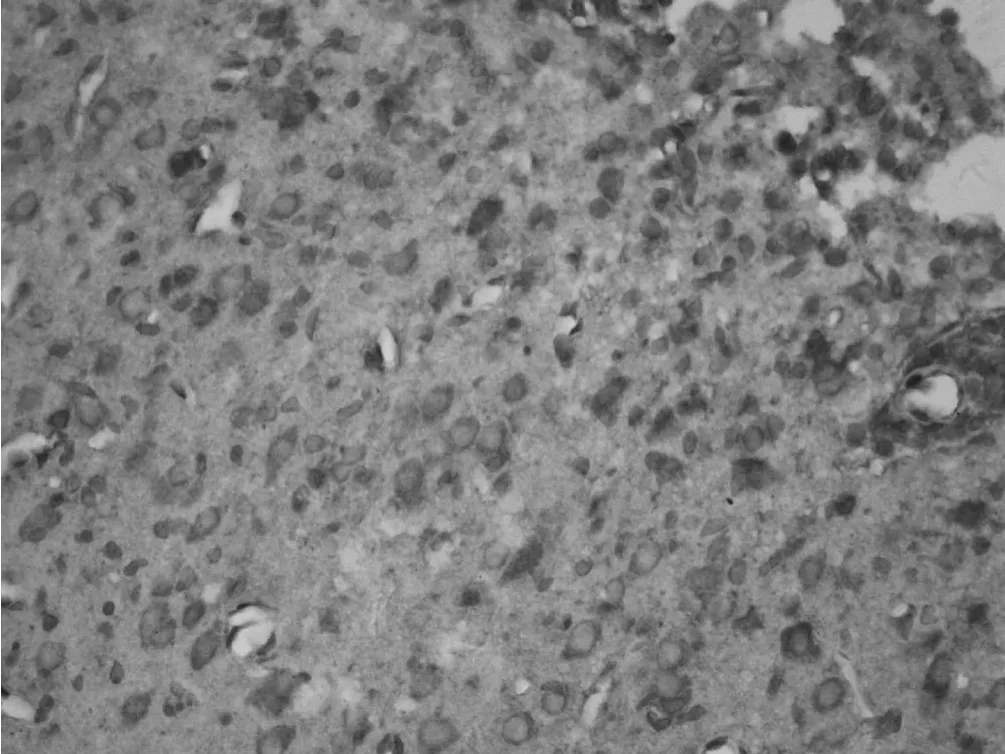
图 3 探索学习组大鼠在术后第 7天时皮质巢蛋白阳性表达情况Figur e 3 Expression of positive nestin neuron in peri-ischemic cortex in learning group at 7 day after MCAO(IHC 400×)
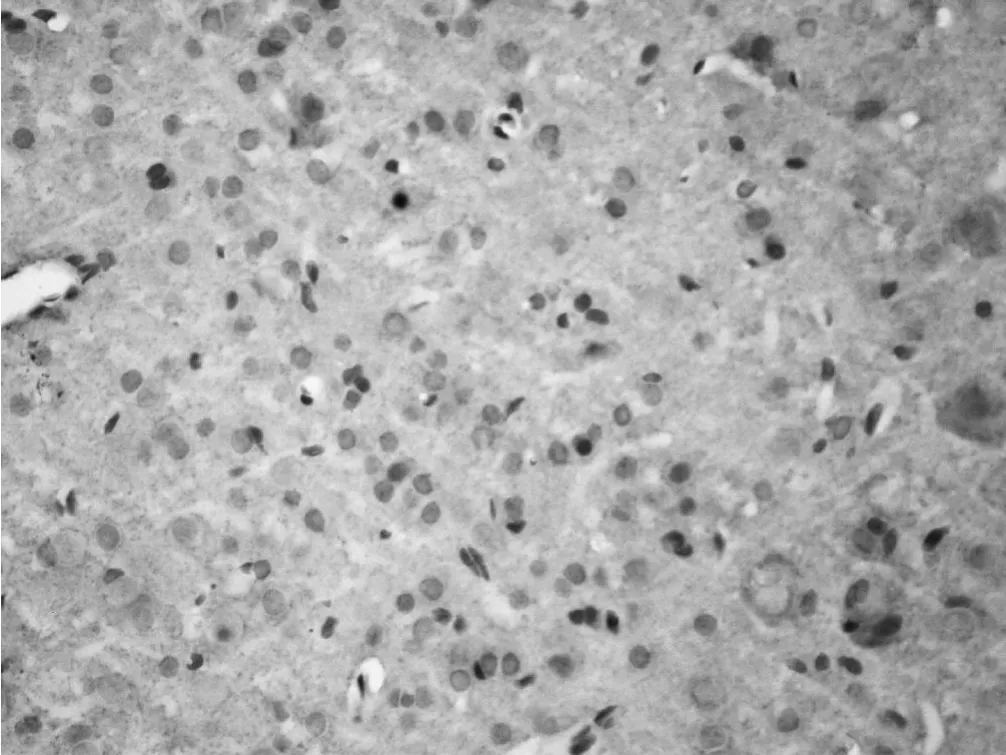
图 4 假手术组大鼠皮质突触素表达情况Figur e 4 Expression of SYN in cortex in sham group(IHC 400×)
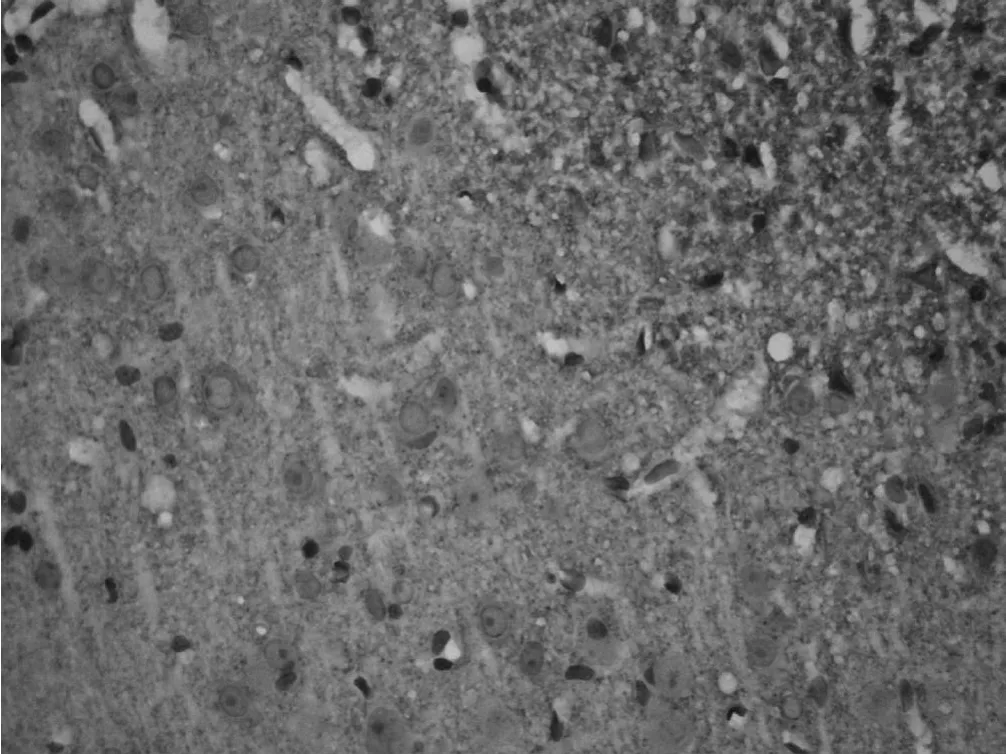
图 5 手术对照组大鼠在术后第 28天时皮质突触素表达情况Figur e 5 Expression of SYN in peri-ischemic cortex in control group at 28 day after MCAO(IHC 400×)

图 6 探索学习组大鼠在术后第 28天时皮质突触素表达情况Figur e 6 Expression of SYN in peri-ischemic cortex in learning group at 28 day after MCAO(IHC 400×)
3 讨论
3.1 探索学习环境对梗死灶周围皮质巢蛋白表达的影响 近十几年来,人们发现在成年脑组织内存在具有多种分化潜能的神经干细胞 (NSC),正常情况下这些细胞处于静息状态,当其所处的微环境发生改变或在外来信号的刺激下,能自我更新,并在一定条件下分化为神经元、星形胶质细胞和少突胶质细胞,参与神经系统的修复,这表明了中枢神经系统 (CNS)具有一定的修复潜能。巢蛋白被认为是 NSC的标志,已广泛用于 NSC的鉴定。巢蛋白是一种新近发现的第六类中间丝蛋白,这种蛋白在哺乳动物胚胎期 CNS的神经前体细胞内大量表达,动物出生后其表达很快降低并消失[4]。但在少数仍保持神经发生功能的部位 (如大脑室管膜下区、嗅球、海马齿状回)仍有表达。这些巢蛋白阳性细胞具有分化和增殖的能力,提示巢蛋白可能参与调节这些细胞的分化与增殖。
随着 Reynolds等[5-6]在成体脑内发现了 NSC,人们愈来愈认识到,NSC在脑损伤修复中起到了非常重要的作用。大量研究表明,NSC在神经营养因子和内环境的调控下可对因缺血损伤而造成的神经功能丧失产生代偿和修复作用[7-8]。在脑缺血的早期即可诱导巢蛋白的表达,从而在脑损伤的修复中发挥主要作用[9]。在损伤最重区域,巢蛋白阳性细胞数目最显著,故巢蛋白可作为 CNS损伤时最早、快速应答的敏感标志物;另一方面,巢蛋白是 NSC的特异性标记蛋白,NSC具有多分化潜能,可以分化成神经元和胶质细胞,并能自我更新。巢蛋白的高表达可能预示了 NSC的激活,从而在脑损伤的修复中发挥重要作用。本实验结果显示,探索学习组大鼠梗死灶周围皮质巢蛋白阳性细胞的数量在造模后第 7、14天明显多于手术对照组,提示探索学习可能通过增强脑梗死大鼠巢蛋白的表达,保护神经元免受缺血缺氧损伤,促进脑功能的重组,从而有利于行为学的恢复。
脑缺血不仅可激活 NSC,使 NSC增殖分化,而且也可使缺血灶周围的某些成熟神经元逆转为 NSC。Li等[10]的实验表明,局灶脑缺血后 6~12 h,缺血灶中心区的星形细胞开始表达,随着时间的推移,缺血灶及周边区小胶质细胞、单核细胞也表达巢蛋白。更为有趣的是,缺血灶附近的一些神经元也呈现巢蛋白阳性。显然这些巢蛋白阳性 NSC并非来源于室管膜下层和齿状回等区,因为他们具有成熟神经元和胶质细胞的特征,提示这种逆向转化的存在。本实验发现,除大脑室管膜下区、海马齿状回外,海马 CA1~4区、缺血灶周围皮质也可见巢蛋白的表达。推测为脑缺血损伤可能刺激脑内成熟的间质细胞或星形胶质细胞返回到 NSC状态,重新分化成受损最严重的细胞类型 (主要是神经元),并迁移至损伤的脑区,从而发挥修复和重排神经元网络的作用[11]。
脑损伤后,巢蛋白的重新表达与神经营养因子的释放有关,而不同环境刺激后,各种神经营养因子的表达水平也不同。因此,探索学习诱导局灶性脑梗死大鼠海马巢蛋白表达,可能是探索学习促进脑梗死后功能恢复的机制之一。
3.2 探索学习环境对梗死灶周围皮质突触素表达的影响 突触素是突触囊泡膜上的一种与突触结构和功能密切相关的钙结合蛋白[12],是突触发生和突触可塑性的重要标志,其参与Ca2+依赖性的神经递质释放,还参与神经元间信息的传递[13]。几乎所有中枢和周围神经系统的突触前终末内均有突触素,它的表达可以反映突触的发生和密度,并且它通过与 Ca2+的结合引起神经递质的释放,还可影响神经信息的传递和加工。它可作为突触前终末的特异性标记物,用来检测突触的密度和分布,是神经元功能状态的标记物之一。突触素膜含量的高低可反映突触神经功能的高低、突触的信息传递、递质释放等神经生物功能正常与否。因其与神经生长、修复、再生和突触重塑密切相关,成为近年来医学研究的热点。
脑功能重组是神经康复的重要理论基础。CNS具有结构和功能重新组织的能力,特别是在损伤后 CNS具有高度的可塑性。轴突出芽、突触形成、突触结构和功能的改变是脑功能重组重要的物质基础。脑缺血损伤可致神经元功能发生不同程度的缺损,随着缺血时间的延长,机体可通过促进突触素的合成作用,引起突触结构和功能两方面的重建。新突触形成的方式有两种:(1)未受损神经元通过轴突绕道投射,树突不寻常分叉等方式进行突触重建;(2)海马齿状回 NSC增殖、迁移和分化,大量新生神经元到达受损区域,与周围神经元形成新的突触连接,以达到代偿目的[14-15],突触前膜功能在缺血后代偿性增加也可使突触素表达升高。本实验结果显示,突触素的免疫活性在第 1天时已有所表达,第 7天时免疫活性增强,可能与未受损神经元的突触重建有关;第 14天免疫活性明显增强,第 28天时最强,可能与 NSC的增殖、迁移、分化有关。
许多研究亦表明学习是影响脑可塑性的主要因素之一,学习会增加突触的数量,改变突触的结构[16]。当大鼠经水迷宫训练获得了空间辨别性学习记忆功能后,在海马内会引起突触数量增多、突触活性区膜面积增加、突触小泡数量和体积增加等一系列形态学变化,这些变化可以认为是空间辨别性学习记忆在大鼠海马结构内引起了突触形态学的可塑性变化[17],而树突棘可能是长期突触可塑性的关键,直接关系到记忆在大脑的存储[18]。探索学习能够向中枢神经提供大量冲动,有利于突触联系的建立,激活神经通路,实现神经功能的重组,从而恢复正常的功能。随着探索学习模型的建立,探索学习的信息不断传入脑内,引起脑内突触的形成或改建。本实验结果表明,探索学习组大鼠梗死灶周围皮质的突触素光密度值在第14、28天较手术对照组为高,有明显差别,说明探索学习与突触素的表达呈正相关性。同时本研究也发现,大鼠经探索学习训练,随着时间的延长,突触素的反应产物明显增多。探索学习引起海马突触素表达增加,这可能是探索学习促进脑梗死后神经损伤恢复和功能重组的机制之一。
1 高俊淑,李阔,李娜,等.探索学习对局灶性脑梗死大鼠梗死灶周围皮质BDNF表达的影响 [J].中国康复医学杂志,2007,22(7):584-585.
2 贾子善,李阔,槐雅萍,等 .不同环境干预对局灶性脑梗死大鼠行为学恢复的影响 [J].中国康复医学杂志,2007,22(7):578-580.
3 Bederson JB,Pitis LH,Tsun M,et al.Rat middle cerebral artery occlusion:evaluation of the model and development of a neurologic examination[J].Stroke,1986,17(3):472-476.
4 Zhigang Jin,Li Liu,Wei Bian,et al.Different transcription factors regulate nestin gene expression during P19 cell neural differentiation and central nervous system development[J].J Biol Chem,2009,20(3):8160-8173.
5 Reynolds BA,Tetzlaff W,Weiss S.A multipotent EGF-responsive striatal embryonic progenitor cell produces neurons and astrocytes[J].JNeurosc,1992,12:4565-4574.
6 Reynolds BA,Seiss S.Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system[J].Science,1992,255(5052):1707-1710.
7 Larsson E,Mandel RJ,Klein RL,et al.Suppression of insult-induced neurogenesis in adultrat brain by brain-derived neurotrophic factor[J].Exp Neurol,2002,177(1):1-8.
8 Teramoto T,Qiu J,Plumier JC,et al.EGF amplifies the replacement of parvalbumin-expressing striatal interneurons after ischemia[J].Clin Invest,2003,111(8):1125-1132.
9 Liu J,Solway K,Messing RO,et al.Increased neurogenesis in the dentate gyrus after transient global ischemia in gerbils[J].Neurosci,1998,18(19):7768-7778.
10 Li Y,Chopp M.Temporal profile of nestin expression after focal cerebral ischemia in adult rat[J].Brain Res,1999,838(1/2):1-10.
11 Abe K.Therapeutic potential of neurotrophic factors and neural stem cells against ischemic brain injury[J].Cereb Blood Flow Metab,2000,20(10):1393-1408.
12 Martinez G,Di Giacomo C,Camazza ML,et al.MAP-2,synaptophysin immunostaining in rat brain and behavioral modifications after cerebral postischemic reperfusion[J].Dev Neurosci,1997,19:457-464.
13 Thiel G.SynapsinⅠ ,SynapsinⅡ and Synaptophysin:marker proteins of synaptic vesicles[J].Brain Pathol,1993,3(1):871.
14 Faverjon S,Silveira DC,Fu DD,et al.Beneficial effects of enriched environment following status epilepticus in immature rats[J].Neurology,2002,59(9):1356-1364.
15 Auvergne R,Lere C,El-bahh B,et al.Delayed kindling epileptogenesis and increased neurogenesis in adult rats housed in an riched environment[J].Brain Res,2002,954(2):277-285.
16 Klintsova AY,Greenough WT.Synaptic plasticity in cortical systems[J].Current Opinion in Neurobiology,1999,9:203-208.
17 宿宝贵,许鹿希 .大鼠海马结构在空间辨别性学习记忆时的突触形态可塑性的定量研究 [J].解剖学研究,2000,22(1):40-42.
18 Leggio MG,Mandolesi L,Federico F,et al.Environmental enrichment promotes improved spatial abilities and enhanced dendritic growth in the rat[J].Behav Brain Res,2005,163(1):78-90.
猜你喜欢
世界科学技术-中医药现代化(2021年10期)2021-03-02
小资CHIC!ELEGANCE(2021年46期)2021-01-11
睿士(2020年11期)2020-11-16
野生动物学报(2020年1期)2020-02-21
现代装饰(2018年5期)2018-05-26
中国高原医学与生物学杂志(2017年4期)2017-03-08
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
中国卫生标准管理(2015年2期)2016-01-14
中国生化药物杂志(2015年4期)2015-07-07
弹箭与制导学报(2015年1期)2015-03-11
