
与狂犬病病毒P 蛋白互作宿主蛋白的筛选与鉴定
2021-01-22 02:56陈贝贝童剑军何川川张雪萍杨勇飞李有文
中国预防兽医学报 2020年12期
陈贝贝,马 海,童剑军,2,苟 涛,何川川,2,张雪萍,高 娜,2,杨勇飞,2,李有文,2*
(1.塔里木大学动物科学学院,新疆 阿拉尔 843300;2.新疆生产建设兵团塔里木畜牧科技重点实验室,新疆 阿拉尔 843300;3.塔里木大学动物科学学院,新疆 阿拉尔 843300)
狂犬病是由狂犬病病毒(Rabies virus,RV)感染引起的急性、直接接触性传染病,以中枢神经系统受到侵害为本病最主要的特征,人与几乎所有温血动物均易感,多因携带RV 的犬猫咬伤或舔舐伤口而感染。临床上有狂躁型或抑郁型两种,特有的临床症状表现为恐水、怕风、咽肌痉挛等,最终因身体部分或全身瘫痪、呼吸衰竭而死,病死率几乎100%。具有非化脓性脑炎的特征性病变,镜下可见神经细胞内的内基氏小体(Negri bodies),可作为诊断该病的重要依据[1]。狂犬病分布范围广泛,世界每年约6 万人死于狂犬病,我国是第二个狂犬病高发国[2-4],加强对该病防治意义重大。
RV 属于弹状病毒科狂犬病病毒属,是一种有囊膜、不分节段、单股负链RNA 病毒,主要编码核蛋白(N)、磷蛋白(P)、基质蛋白(M)、糖蛋白(G)和聚合酶蛋白(L)等5 个重要的结构蛋白[5]。P 蛋白是多功能蛋白,被认为是RV 的“外交官”[6]。一方面,P 蛋白可以与病毒的多种蛋白相互作用,保证病毒正常复制。例如:P 蛋白可以自身相互作用形成二聚体[7];也可与L 蛋白结合对病毒转录和复制起至关重要的作用[8];还可以与N-RNA 复合体结合参与病毒的转录[9];作为N 蛋白的分子伴侣引导N蛋白正确折叠[10]。另一方面,P 蛋白又可以与宿主细胞的多种蛋白相互作用,抵抗宿主的免疫防御。例如:P 蛋白可以与干扰素通路相关蛋白PML、STAT 相互作用阻止干扰素对病毒复制的抑制作用[11];还可以与宿主动力蛋白轻链LC8 相互作用,促进病毒增殖[12]。因此,对P 蛋白的研究有利于对RV 致病机理的了解和认识。
噬菌体展示技术(Phage display technology)是一种研究蛋白互作的传统方法,该方法先将组织细胞基因克隆到噬菌体中,构建噬菌体文库,并使基因编码的蛋白表达在噬菌体表面,与目的蛋白结合,通过洗脱筛选和测序得到与宿主互作蛋白的基因信息[13]。为了获得更多与RV P 蛋白相互作用的宿主细胞蛋白,本研究经原核系统表达了P 蛋白,并利用噬菌体展示技术从人脑组织cDNA 噬菌体文库中筛选与P 蛋白互作的蛋白,为进一步研究P 蛋白、了解RV 的致病机制奠定基础。
1 材料与方法
1.1 主要实验材料RV SAD-L16 株由华中农业大学农业微生物国家重点实验室保存;含RV P 基因的质粒pEGF-P 由华中农业大学彭贵青教授惠赠;人脑组织T7 噬菌体cDNA 文库、噬菌体宿主菌5403购自Novagen 公司;大肠杆菌DH5α、BL21、宿主菌、原核表达载体pET42b 均由新疆生产建设兵团塔里木畜牧科技重点实验室保存。
PrimeSTAR HS DNA Polymerase、dNTP、T4 DNA连接酶均购自TaKaRa 公司;Nde I、Xho I 购自Ther⁃mo 公司;DNA 纯化试剂盒购自美国Omega 公司;琼脂糖凝胶回收试剂盒、质粒小提试剂盒购自天根生化科技(北京)公司;30%丙烯酰胺、显色剂A/B 购自碧云天公司;蛋白纯化试剂盒购自北京康为世纪生物科技有限公司;T7 噬菌体抗体购自Novagen 公司;P 蛋白单克隆抗体(MAb)由华中农业大学赵凌教授惠赠;HRP 标记的羊抗鼠多克隆抗体购自博士德生物有限公司。
1.2 引物的设计与合成根据RV SAD-L16 株的P基 因 序 列(EU182347), 设 计 引 物SADL16-P1/SADL16-P2 PCR 扩增P 基因,并在其上下游引物的5'端分别引入Nde I 和Xho I 酶切位点;根据T7 噬菌体文库载体序列,设计T7 10-3b 株引物T7 10-3b P1/T7 10-3b P2,以扩增筛选出的互作蛋白基因(表1)。引物均由武汉天一辉远生物有限公司合成。
1.3 原核表达载体的构建与鉴定以pEGFP-P 质粒为模板,利用SAD-L16-P1/P2 引物PCR 扩增P 基因,经Nde I 和Xho I 双酶切后插入pET42b 质粒构建P 基因的原核表达载体,载体经Nde I 和Xho I 双酶切和测序鉴定,正确的表达载体命名为pET42b-P。根据测序结果利用软件DNAMAN 和DNAStar 对P 蛋白二级结构进行分析。
1.4 P 蛋白的原核表达、鉴定及纯化pET42b-P 转化大肠杆菌BL21 感受态细胞,37 ℃振摇培养至OD600nm为0.6 时,加终浓度为1 mmol/L 的IPTG 诱导表达4 h~6 h 后,离心弃上清收集菌体,超声破碎,离心后取上清和沉淀分别经SDS-PAGE 凝胶电泳检测P 蛋白的表达,以P 蛋白MAb(1∶1 000)为一抗,HRP 标记的羊抗鼠多克隆抗体(1∶1 000)为二抗,经western blot 鉴定。表达蛋白经蛋白纯化试剂盒纯化(按试剂盒说明书操作)后通过SDS-PAGE 检测纯化效果,并利用核酸蛋白仪测定其浓度。

表1 引物信息Table 1 Primer information
1.5 噬菌体展示技术筛选与P 蛋白互作的宿主蛋白参考文献[14]筛选与P 蛋白互作的宿主蛋白:首先取纯化的P 蛋白(10 mg/L)包被聚苯乙烯微量反应板,并用5%的脱脂奶粉封闭;然后用T7 噬菌体宿主菌5403 培养扩增人脑组织T7 噬菌体cDNA 文库后,将文库加入包被了P 蛋白的微量反应板,常温放置2 h 后弃去文库并彻底清洗反应孔以除去没有作用的噬菌体,用解析缓冲液洗脱并收集洗脱液为第1 轮筛选文库。取部分文库采用涂板计数噬菌斑的方法测定其库容量,剩余文库接种5403 宿主菌培养扩增。取第1 轮筛选文库加入包被P 蛋白的聚苯乙烯微量反应板,同前法进行第2 轮筛选。如此经5 轮筛选后挑取全部噬斑分别作为模板,利用引物T7 10-3b P1/P2 进行PCR 扩增,PCR 产物由武汉天一辉远生物有限公司测序。测定序列用NCBI 中BLAST 软件进行相应基因序列对比分析,确定可能与P 蛋白互作的候选宿主蛋白。
1.6 候选互作宿主蛋白的ELISA 验证为了进一步验证筛选的互作蛋白及其与P 蛋白作用的强弱,将纯化的P 蛋白(10 mg/L)包被聚苯乙烯微量反应板,分别以100 μL/孔加入1∶10 的候选互作宿主蛋白噬菌体文库为一抗,同时设PBS、T7 噬菌体原库及宿主菌5403 为对照。以100 μL/孔T7 噬菌体抗体(1∶10 000)为二抗,通过ELISA 检测与P 蛋白互作的候选宿主蛋白,及其与P 蛋白的反应性。
2 结 果
2.1 原核表达载体的构建与鉴定结果构建的P 基因原核表达载体经Nde I/Xho I 双酶切鉴定,结果显示得到了与预期相符的载体和目的基因两条片段(图1);测序结果表明P 基因全长894 bp,与参考株ERN.NS P 基因的同源性达99.44%,表明pET42b-P表达载体构建正确。

图1 表达载体pET42b-P 的双酶切鉴定结果Fig. 1 Identification of pET42b-P by endonuclease digestion
2.2 P 蛋白的二级结构分析通过DNAStar 软件预测,RV SAD-L16 株P 蛋白由297 个氨基酸构成,其二级结构以α 螺旋结构及无规则卷曲交错形成,β转角较少,P 蛋白亲水性很强,柔韧性和抗原性均较好(图2)。

图2 P 蛋白的二级结构分析Fig. 2 Secondary structure prediction of P protein
2.3 P 蛋白的表达与鉴定将表达载体pET42b-P转化BL21 并经IPTG 诱导后,SDS-PAGE 结果显示,在上清和沉淀中均有大小约为38 ku 的目的条带,均与理论P 蛋白大小相符,且上清中蛋白的量更多。进一步利用P 蛋白MAb 经western blot 检测,结果显示,在约38 ku 处出现特异性条带(图3A)。表明经原核系统表达了P 蛋白且P 蛋白以可溶形式表达。将表达的P 蛋白利用镍层析法纯化,经SDSPAGE 检测结果显示,蛋白纯化效果较好(图3B)。利用核酸蛋白测定仪测定蛋白含量为230 mg/L。
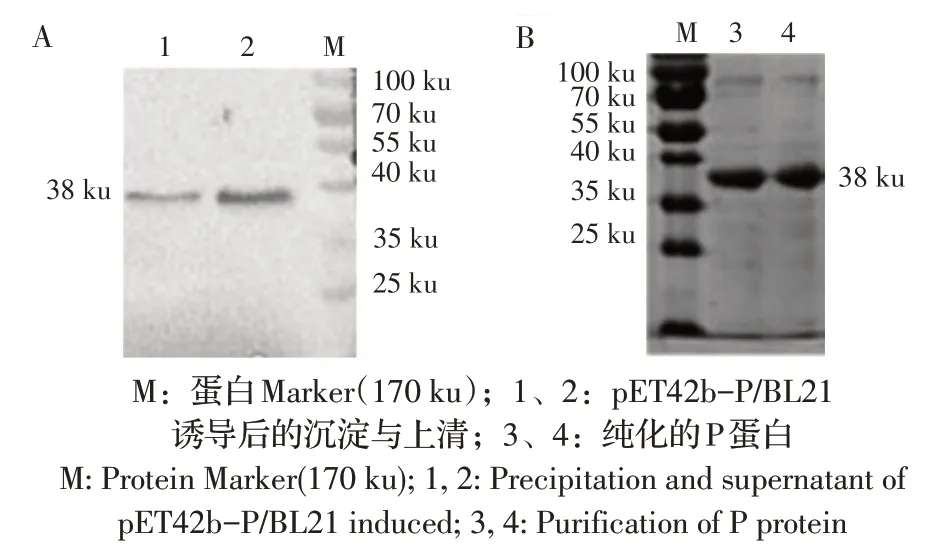
图3 P 蛋白表达的western blot(A)及其纯化效果的SDS-PAGE(B)鉴定结果Fig. 3 Identification of P protein expression by western blot (A) and its purification by SDS-PAGE(B)
2.4 与P 蛋白互作宿主蛋白的筛选结果采用噬菌体展示技术经过5 轮筛选共挑取噬斑31 个,并以其为模板,采用引物T7 10-3b P1/P2 经PCR 鉴定。结果显示,得到24 种扩增产物(图4),全部测序后经NCBI 的BlAST 比对,经DNAMAN 软件分析合并相同基因序列后共得到18 种候选蛋白。根据噬菌体文库的载体序列,并且利用DNAstar 分析筛选出的蛋白基因序列的表达框是否与载体的表达框相匹配,最终确定能正确表达的候选互作蛋白有5 种(表2)。

图4 筛选的与P 蛋白互作蛋白基因的PCR 鉴定结果Fig. 4 Identification of the selected interacting protein gene by PCR
2.5 候选互作宿主蛋白的ELISA 验证结果筛选的5 种候选互作宿主蛋白经ELISA 检测结果显示,筛选蛋白的噬菌体文库的OD630nm值均高于0.7,甚至超过1.0,而PBS、空宿主菌、未经筛选的噬菌体文库等阴性对照的OD630nm值均在0.1 以下(OD630nm值≥0.4 表示候选宿主蛋白与RV P 蛋白存在相互作用,图5)。表明5 个筛选的候选互作宿主蛋白均有可能与P 蛋白相互作用,其中以泛素特异性肽酶54 和核糖体蛋白L15 的作用更强,作为与P 蛋白互作蛋白的可能性较大。

表2 最终筛选的与P 蛋白互作的5 种候选宿主蛋白信息Table 2 Selected candidates proteins interacting with P protein
3 讨 论
RV P 蛋白是一种重要的结构蛋白,在病毒的增殖和感染宿主过程中起着重要的辅助作用,其功能非常活跃。但P 蛋白很多功能还未被发现,其结构还未被解析,狂犬病的致病机制也不十分清楚,因此对P 蛋白进行深入研究,寻找与P 蛋白互作的宿主蛋白以及这种互作对病毒复制的影响机制,将有助于抗病毒新靶标药物的研发。
本研究利用pET42b 原核表达载体经PCR 扩增了RVSAD-L16 株的P 基因并测序,序列分析发现其由894 个核苷酸组成,与RVERN.NS 参考株P 基因的同源性为99.44%。P 基因比对结果显示,张强等对我国不同地区不同年代分离的4 株RV 与参考株同源性最低为83.6%,最高为99.8%[15],表明P 基因变异较大。利用DNAStar 软件预测P 基因编码297 个氨基酸,分子量大小约35.6 ku,但本实验表达的P 蛋白经SDS-PAGE 结果显示其约为38 ku,与预测有差异,但与李有文等表达的P 蛋白的结果一致[16]。根据章正瑛等分析,由于凝胶特性与蛋白带电等多种因素的影响,SDS-PAGE 电泳结果与预测可能会有1 ku~5 ku 的差异,属正常现象[17]。对其二级结构预测并分析发现其亲水性非常强,也具有较好的柔性。

图5 与P 蛋白互作宿主蛋白的ELISA 验证结果Fig. 5 ELISA results of five candidate proteins interacting with P proteins
噬菌体展示技术是研究蛋白质相互作用的传统手段,其以成本低、操作简单而广泛应用于治疗性MAb 的开发[18]、多肽类药物研究[19-20]、疫苗研发[21]以及食品安全分析[22]。目前主要有丝状噬菌体、T7 噬菌体、T4 噬菌体与λ 噬菌体等展示系统,其中丝状噬菌体展示系统应用最为广泛[23]。从理论上讲,只要是与病毒靶蛋白存在可能互作的宿主蛋白,利用噬菌体展示技术就可以将其筛选出来,不会被遗漏,但实际操作中可能会受多种因素的限制而不能将其全部筛选出来。本研究利用原核系统表达并纯化获得RV P 蛋白,并利用噬菌体展示技术从商品化的人脑组织T7 噬菌体文库经5 轮筛选获得的全部噬斑经PCR 鉴定、测序分析和BLAST 比对后,筛选出18 种与P 蛋白存在相互作用的候选宿主蛋白。进一步将各候选蛋白的基因序列与噬菌体载体的表达框对比分析以确定其是否正确表达,最终确定能正确表达的候选互作宿主蛋白只有5 种。
利用噬菌体展示技术筛选与病毒蛋白互作的宿主蛋白,其结果存在一定的假阳性或假阴性[24-25],因此,筛选出来的宿主蛋白是否是真正的互作蛋白还需进一步的实验验证。常用的验证方法有荧光定量PCR、Co-IP、Pull-down、细胞共定位和ELISA等[13]。本研究采用ELISA 方法检测经筛选得到的5种候选宿主蛋白与P 蛋白的相互作用,反向证明候选宿主蛋白是否与P 蛋白存在相互作用,结果表明泛素特异性肽酶54 和核糖体蛋白L15 与P 蛋白互作的可能性更大。根据这两种蛋白可能的生物学功能,推测P 蛋白与泛素特异性肽酶54 互作可能会影响宿主蛋白的去泛素化,从而影响细胞的正常代谢、免疫调节而利于病毒感染;P 蛋白与核糖体蛋白L15 的互作可能会引起宿主蛋白的合成异常,从而有益于病毒复制而致病等。
本研究利用噬菌体展示技术从人脑组织T7 噬菌体文库中筛选与RV P 蛋白互作的宿主蛋白,不仅为进一步了解P 蛋白的生物学功能奠定基础,也为RV 其他蛋白生物学特性的研究提供思路。
猜你喜欢
湘潮(上半月)(2022年7期)2022-12-06
中国动物保健(2022年2期)2022-05-05
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年4期)2021-11-12
猪业科学(2021年3期)2021-05-21
幽默大师(2020年10期)2020-11-10
科学24小时(2020年4期)2020-05-14
中华诗词(2019年1期)2019-11-14
科学大观园(2019年23期)2019-09-10
安徽医科大学学报(2015年9期)2015-12-16
