
胸膜肺炎放线杆菌ApxⅠ、ApxⅢ和ApxⅣ的截短表达及免疫反应性鉴定
2021-05-28 04:14于栋祝可心安家慧叶洋李玉峰
畜牧与兽医 2021年6期
于栋,祝可心,安家慧,叶洋,李玉峰
(南京农业大学动物医学院/农业农村部动物细菌学重点实验室,江苏 南京 210095)
猪传染性胸膜肺炎是猪的一种呼吸系统急性接触性传染病,临床症状包括高烧、咳嗽和流鼻涕[1]。该病各年龄阶段的猪群均易感,发病率在10%以上,最急性型死亡率可达80%~100%,从急性和慢性感染中康复的猪可以持续排出病原体而没有临床症状,因此难以控制和根除[2-4]。自1957年发现以来,猪传染性胸膜肺炎对各国生猪产业造成了巨大的经济损失[5]。胸膜肺炎放线杆菌(Actinobacilluspleuropneumoniae,APP)属于巴氏杆菌科、放线杆菌属,是一种革兰阴性短杆状细菌,是猪传染性猪胸膜肺炎的重要致病因子[1]。APP共有15种血清型,不同血清型间交叉保护性低[5-6]。APP能够产生多种毒力因子,包括溶血外毒素(Actinobacilluspleuropneumoniae-RTX-toxins,Apx)、外膜蛋白、脂多糖、转铁结合蛋白、荚膜、菌毛等[7],其中 Apx被认为是APP最重要的毒力因子。目前已经发现的Apx有ApxⅠ、ApxⅡ、ApxⅢ和ApxⅣ[8],其中ApxⅠ有较强的溶血活性;ApxⅡ溶血活性较弱[9];ApxⅢ有强细胞毒性,不具有溶血活性[10];ApxⅣ由所有APP血清型在感染猪体中产生,但在体外培养条件下不产生[2],由大肠杆菌表达的产物有较弱溶血活性[11]。与此同时,Apx也是APP主要的免疫原,在针对APP的各种疫苗中,使用ApxⅠ、ApxⅡ和ApxⅢ的重组蛋白作为疫苗抗原制备的亚单位疫苗在保护性方面显示出良好的功效:马艳芳[12]研究提取ApxⅠ、ApxⅡ、ApxⅢ毒素,发现其在猪传染性胸膜肺炎的免疫保护过程中具有一定作用;周家强[9]将ApxⅡ A蛋白纯化后免疫小鼠,三免后发现其抗体效价明显提升,并且通过动物试验证明ApxⅡ A蛋白在一定程度内能够增强灭活苗的免疫效果;王方昆等[13]研究发现ApxⅣ N端有良好的免疫原性;万玉萌等[14]表达了ApxⅠ、ApxⅡ、ApxⅢ和外膜蛋白Omp2,并证明重组蛋白具有良好的免疫反应性。APP血清学诊断方法的建立也常以Apx为抗原:Myunghwan等[2]开发的ELISA方法可以评估动物体内针对ApxⅠ,ApxⅡ和ApxⅢ每种毒素的特异性抗体水平;李凤梅等[5]研究发现以ApxⅣ-ELISA检测APP具有一定的可行性,且具有特异性良好、准确性较高、重复性较好的特点。
目前,国内相关研究中制备的Apx毒素蛋白大多以包涵体形式存在,免疫原性较差。本研究选择APP的ApxⅠ、ApxⅢ、ApxⅣ亲水区进行截短表达,以期提高重组蛋白在上清的表达量,为后续亚单位疫苗的制备以及建立具有较好特异性的ELISA检测方法提供参考。
1 材料与方法
1.1 菌株、质粒
大肠杆菌DH5α、Rosetta(DE3),APP血清2型和5型等由本实验室保存;原核表达载体pColdI、pET-32a购自TaKaRa(大连)生物技术有限公司。
1.2 试剂
T4 DNA连接酶,购于TaKaRa(大连)公司;DNA凝胶回收盒、异丙基硫代半乳糖苷(IPTG)、琼脂糖、小鼠抗6×His组氨酸单克隆抗体(mAb),购于南京天为公司;DNA Marker DL5000、2× Phanta Mix,购于诺唯赞公司;DNA Marker DL2000,购于擎科生物科技有限公司;辣根过氧化物酶(HRP)标记的羊抗鼠IgG,购于南京生兴生物技术有限公司;蛋白分子质量标准品、限制性核酸内切酶XhoⅠ、HindⅢ和SacⅠ,购于赛默飞世尔科技(中国)公司;细菌组DNA提取盒,购于天根公司;质粒提取试剂盒,购自Biomiga;其余试剂为进口/国产分析纯。
1.3 截矩基因片段tApxⅠ、tApxⅢ、tApxⅣ的扩增
参照TIANamp Bacteria DNA Kit说明书,取3 mL过夜转接的新鲜APP 2型和5型菌液提取总DNA。根据GenBank中收录的APP ApxⅠ、ApxⅢ、ApxⅣ基因序列设计引物,并在ApxⅠ、ApxⅢ、ApxⅣ 3对引物的上、下游分别对应引入XhoⅠ和HindⅢ的2个酶切位点,在ApxⅢ引物的上、下游分别对应引入SacⅠ和XhoⅠ的2个酶切位点。引物合成由英潍捷基公司完成(序列见表1)。
以APP菌液提取的总DNA为模板,用上述引物扩增tApxⅠ、tApxⅢ、tApxⅣ基因。PCR反应体系如下:2×Phanta Mix 12.5 μL,上游引物1.0 μL,下游引物1.0 μL,DNA模板2.0 μL,补水至25 μL。PCR扩增条件如下:95 ℃预变性5 min;95 ℃变性30 s,退火30 s,72 ℃延伸2 min,进行35个反应循环;72 ℃延伸10 min。
1.4 原核表达质粒的构建与鉴定
PCR产物经1%琼脂糖核酸凝胶电泳进行初步鉴定,参照凝胶回收试剂盒说明进行片段回收,将含有酶切位点的目的基因片段和pColdI空表达载体同时进行限制性核酸内切酶双酶切,上述2种产物分别经10 g/L琼脂糖凝胶核酸电泳,根据目的片段大小分别回收产物后进行连接,上述连接产物转化进入克隆感受态DH5α细胞内,经37 ℃ 摇床200 r/min培养1 h后涂布于LB固体培养基(pColdI为Ampr)37 ℃过夜培养。经PCR鉴定,挑取疑似阳性的单克隆菌落,接种于LB液体培养基(内含有Ampr)培养12~16 h后提取质粒,对重组质粒进行双酶切鉴定,鉴定结果为阳性后,送至通用公司测序,将所得的序列提交到NCBI网站上GenBank进行序列比对,确认正确的重组质粒命名为pColdI-tApxⅠ、pColdI-tApxⅢ、pColdI-tApxⅣ。

表1 引物设计
1.5 重组质粒的诱导表达及蛋白可溶性分析
将鉴定正确的重组质粒转化进入Rosetta(DE3)感受态细胞中,涂布于LB固体培养基(pColdI、pET-32a为Ampr)置37 ℃过夜培养。从LB固体培养基上挑取阳转单克隆菌落,接种至LB液体培养基(pColdI、pET-32a为Ampr),常规培养至菌液OD600范围为0.4~0.6,加入IPTG使终浓度达到1 mmol·L-1,pColdI-tApxⅠ、pColdI-tApxⅢ、pColdI-tApxⅣ以160 r/min 15 ℃的条件诱导24 h后收集1 mL菌液,pET-32a- tApxⅠ以160 r/min 30 ℃的条件诱导8 h后收集1 mL菌液,同时收集诱导的pColdI、pET-32a空载体菌液做阴性对照。收集的菌液经pH=7.4的PBS缓冲液洗涤2次后,以80 μL PBS缓冲液重悬并加入20 μL 5×SDS Loading buffer振荡混匀瞬时离心煮沸5~10 min处理,经SDS-PAGE分析,考马斯亮蓝染色分析。获得诱导后的重组蛋白,收集诱导表达的全菌以pH=7.4的PBS缓冲液洗涤2次并重悬,超声波破碎裂解细胞,分别对上清和沉淀进行收集及处理,经12.5%凝胶SDS-PAGE检测,判断表达产物存在形式。
1.6 重组蛋白的Western blot鉴定及反应原性分析
将重组蛋白rtApxⅠ、rtApxⅢ、rtApxⅣ的表达产物处理后的上清经SDS-PAGE分离,将电泳后产物通过半干法转印至硝酸纤维素膜,将6×His单抗/猪多抗作为一抗,HRP-羊抗鼠/抗猪IgG作为二抗,进行Western blot试验。
1.7 重组蛋白的纯化
将100 mL经IPTG诱导的菌液用PBS缓冲液洗涤2次后以结合缓冲液重悬进行超声裂解,13 000 r/min离心10 min后取上清用0.22 μm滤器过滤后作为蛋白样品。参照Ni-NTA说明书,先将10倍柱体积结合缓冲液通过镍柱,然后加入蛋白样品,之后用洗涤缓冲液洗涤树脂,至流穿液中溶质含量保持稳定,最后以5倍柱体积洗脱缓冲液洗脱树脂,收集纯化样品。
2 结果与分析
2.1 重组质粒的PCR及双酶切鉴定
挑取抗性平板上的单克隆菌落培养提取质粒,进行PCR扩增及核酸凝胶电泳分别出现大小为1 713、1 350、1 467 bp左右的条带(图1A),与预期大小一致。重组质粒pColdI-tApxⅢ经SacⅠ和XhoⅠ双酶切,得到1 350 bp的基因片段和4 400 bp左右的载体片段;pColdI-tApxⅠ、pColdI-tApxⅣ经HindⅢ和XhoⅠ双酶切分别获得1 713、1 467 bp左右的基因片段及4 400 bp的载体片段(图1B)。测序结果正确,表明已成功构建了重组表达质粒pColdI-tApxⅠ、pColdI-tApxⅢ、pColdI-tApxⅣ。
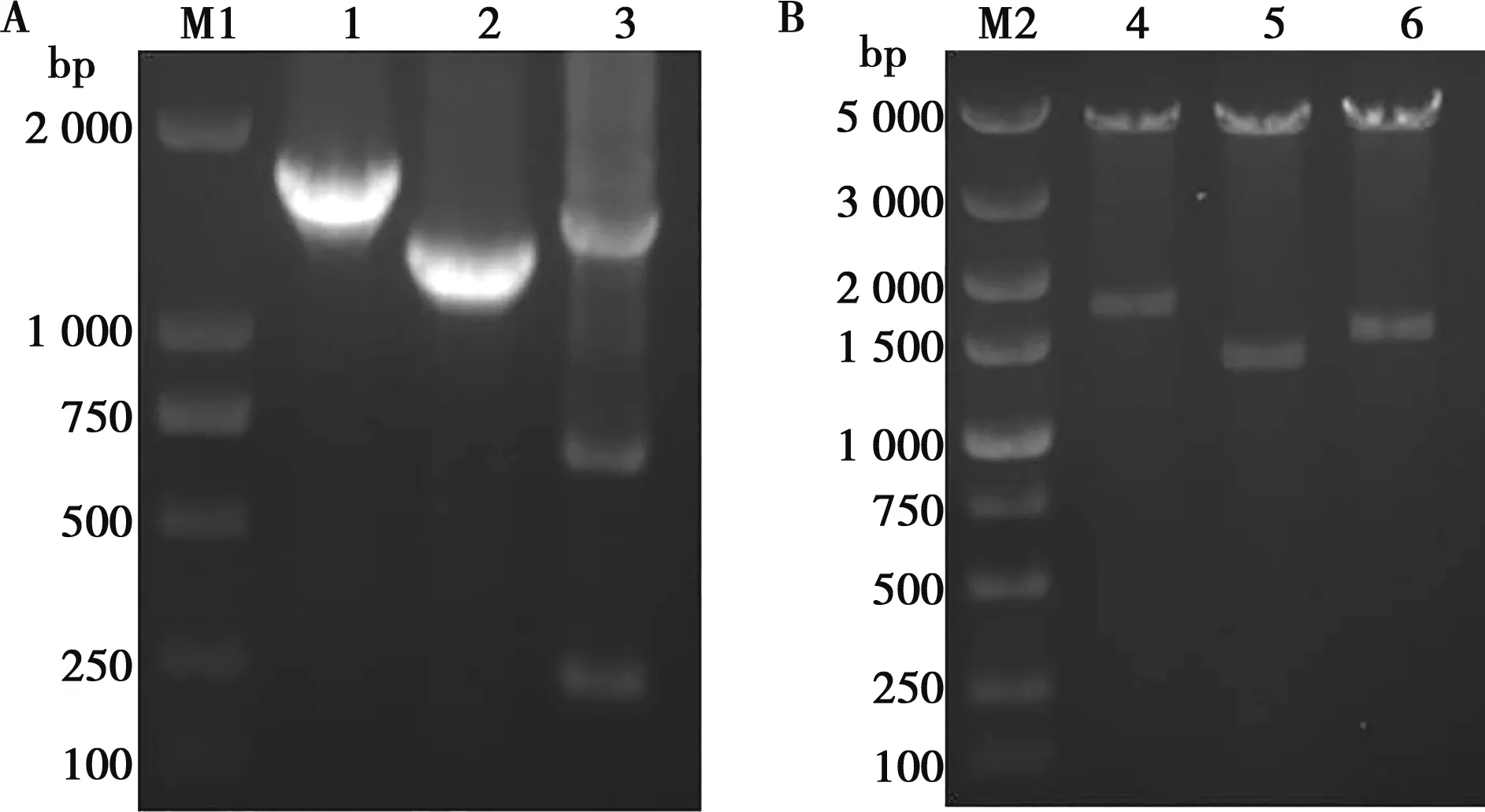
M1.DNA分子质量标准DL2000;M2.DNA分子质量标准DL5000;1.tApxⅠ基因;2.tApxⅢ基因;3.tApxⅣ基因;4.pColdI-tApxⅠ双酶切;5.pColdI-tApxⅢ双酶切;6.pColdI-tApxⅣ双酶切
2.2 重组蛋白的诱导表达
将重组质粒pColdI-tApxⅠ、pColdI-tApxⅢ、pColdI-tApxⅣ转化至Rosetta(DE3)诱导表达,进行SDS-PAGE分析,在63、50和54 kDa左右的位置出现目的条带,与预期相符。确定3种重组蛋白表达以后,将诱导后的细菌全菌经过洗涤,超声波破碎裂解离心,对经过离心的上清液进行凝胶SDS-PAGE分析。pColdI-tApxⅠ、pColdI-tApxⅢ、pColdI-tApxⅣ在上清均表达(图2A、2B)。
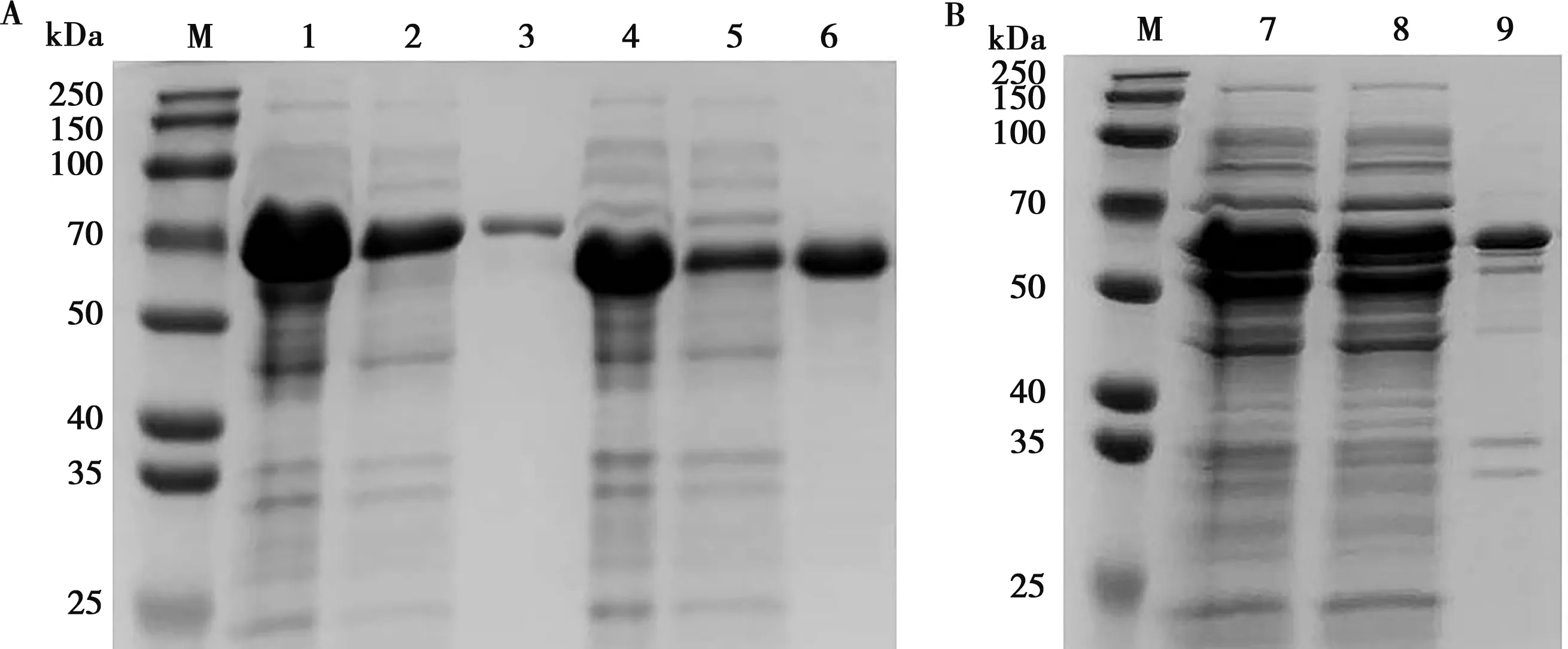
M.预染蛋白分子质量标准;1.Rosetta/pColdI-tApxⅠ诱导后全菌;2.Rosetta/pColdI-tApxⅠ超声破碎后上清;3.Rosetta/pColdI-tApxⅠ超声破碎后沉淀;4.Rosetta/pColdI-tApxⅢ诱导后全菌;5.Rosetta/pColdI-tApxⅢ超声破碎后上清;6.Rosetta/pColdI-tApxⅢ超声破碎后沉淀;7.Rosetta/pColdI-tApxⅣ诱导后全菌;8.Rosetta/pColdI-tApxⅣ超声破碎后上清;9.Rosetta/pColdI-tApxⅣ超声破碎后沉淀
2.3 重组蛋白的Western blot鉴定
对含有重组表达质粒pColdI-tApxⅠ、pColdI-tApxⅢ、pColdI-tApxⅣ的重组大肠杆菌Rosetta诱导表达进行可溶性分析,发现目的蛋白在上清与包涵体中均存在。诱导后的上清液加入上样缓冲液煮沸处理,经SDS-PAGE分析,以鼠抗His-IgG为一抗,HRP-羊抗鼠IgG为二抗进行Western blot分析,在63、50和54 kDa相应位置出现特异性抗原抗体结合带(图3A)。同时,以阳性猪血清作为一抗进行孵育,然后以酶标猪二抗进行孵育后染色,结果证明表达的可溶性重组蛋白rtApxⅠ、rtApxⅢ、rtApxⅣ均与APP天然抗体发生特异性反应(图3B、3C)。
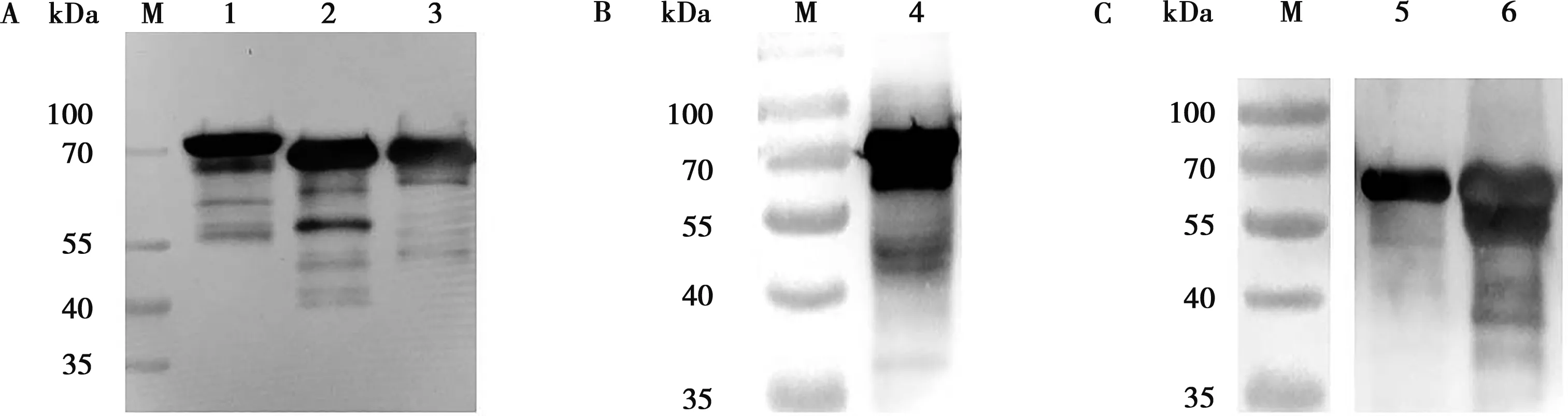
M.预染蛋白分子质量标准;1.超声破碎后Rosetta/pColdI-tApxⅠ上清;2.超声破碎后Rosetta/pColdI-tApxⅢ上清;3.超声破碎后Rosetta/pColdI-tApxⅣ上清;4.超声破碎后Rosetta/pColdI-tApxⅠ上清;5.超声破碎后Rosetta/pColdI-tApxⅢ上清;6.超声破碎后Rosetta/pColdI-tApxⅣ上清
2.4 重组表达蛋白的纯化
SDS-PAGE结果表明,使用Ni-NTA亲和层析纯化,在0 mmol/L结合缓冲液、60 mmol/L 洗涤缓冲液的条件下能够有效纯化ApxⅠ(图4A);在30 mmol/L 结合缓冲液、100 mmol/L洗涤缓冲液的条件下能够有效纯化ApxⅢ(图4B);在20 mmol/L 结合缓冲液、60 mmol/L洗涤缓冲液的条件下能够有效纯化Ⅳ(图4C)。
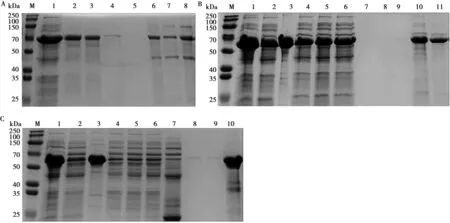
A.重组蛋白rtApxⅠ纯化的SDS-PAGE分析:M.预染蛋白分子质量标准;1.超声破碎后上清;2~3.流穿液;4~5.洗涤缓冲液;6~8.纯化后蛋白
3 讨论
由于包涵体需要经过复杂的变性溶解和复性折叠过程,导致目的蛋白复性率低、损失量大,而可溶性蛋白表达则可以很好地避免这一问题。为了实现目的蛋白的可溶性表达,本试验中对ApxⅠ、ApxⅢ、ApxⅣ的表达曾尝试使用pColdI载体、pET-32a载体,更换宿主菌Rosetta,使用4种带有伴侣蛋白的BL21质粒(PG-KJE8、pGro7、pKJE7、pG-Tf2),并且对诱导温度、转速、时长、IPTG浓度以及诱导前的菌液浓度等条件进行优化,最终实现了pColdI-tApxⅠ、pColdI-tApxⅢ、pColdI-tApxⅣ的可溶性表达。pET-32a载体虽然带有融合蛋白有利于实现可溶性表达,但重组蛋白用于后续试验时需去除载体的融合蛋白,且用pET-32a载体的表达效果不如pColdI载体,故选择pColdI载体进行蛋白的可溶性表达。重组蛋白带有His标签,可使用Ni-NTA进行纯化,通过优化结合液、洗涤液、洗脱液中的咪唑浓度,从而获得了纯化效果较好的rtApxⅠ、rtApxⅢ、rtApxⅣ重组蛋白。
目前,我国主要通过接种全菌灭活疫苗来预防猪传染性胸膜肺炎,但由于APP血清型较多,不同血清型之间交叉保护性低,常规疫苗的使用已经无法达到有效预防该疾病的目的。Park等[15]研究曾表明使用重组Apx毒素的抗APP亚单位疫苗对不同血清型具有良好的交叉保护作用,因此当前许多可用的疫苗都将多个血清型共有的Apx作为其关键抗原[16]。刘建杰等[17]利用大肠杆菌表达APP的分泌毒素ApxⅠ、ApxⅡ、ApxⅢ,提取表达产物包涵体,再加入APP 7型菌制成亚单位菌苗,结果发现用APP 1型和APP 7型菌株进行攻毒后其保护力分别为83.3%和91.7%。邵美丽等[18]用重组蛋白ApxⅠ、ApxⅡ、ApxⅢ、Omp组合作为免疫原免疫小鼠,结果发现,接种4组重组蛋白小鼠的特异性抗体水平较高,且该组用APP 1型菌和APP 2型菌进行攻毒后的保护率也优于其他组。Park等[15]研究中开发的由ApxⅠA和ApxⅡA片段组成的二价疫苗显示出针对APP血清1型、2型菌株的交叉保护作用。本研究在实验室可溶性表达蛋白的基础上,成功对tApxⅠ、tApxⅢ进行了可溶性表达,期望这2种重组蛋白可以共同作为有效抗原成分,从而制备出具有较好免疫保护效果的亚单位疫苗。
ELISA检测方法操作简便、快速,且具有良好的敏感性,是常用的血清学诊断方法。ApxⅣ只在动物体内表达且仅由APP产生,根据这一特点建立的ApxⅣ-ELISA可有效区分APP灭活疫苗或亚单位疫苗免疫猪与自然感染猪[19],并能与其他的猪呼吸道疾病病原如猪链球菌、副猪嗜血杆菌等进行鉴别诊断[13,19]。González 等[20]利用ELISA方法检测了在试验条件下接种APP的猪的血清和口腔液中的ApxⅣ特异性抗体,结果发现接种APP血清1型和7型的猪的ApxⅣ血清和口腔液的抗体反应与LPS抗体反应模式相近。市场上在售的APP-ELISA检测试剂盒中,国外的试剂盒价格昂贵,国内的试剂盒采用的是间接ELISA检测方法,特异性较差。本研究将ApxⅣ表达在上清,后期可以制备ApxⅣ特异性单克隆抗体并将其应用于高效、灵敏地阻断ELISA方法的建立,以期研制出成本低廉且具有较好特异性的ApxⅣ-ELISA诊断试剂盒。
综上所述,本研究在实验室前期对ApxⅡ和Omp2蛋白研究的基础上,通过条件优化使tApxⅠ、tApxⅢ、rtApxⅣ重组蛋白表达在上清并成功纯化,为研制具有较好交叉保护性的亚单位疫苗以及建立有效区分APP灭活疫苗或亚单位疫苗免疫猪与自然感染猪的ELISA检测方法奠定了物质基础。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
中国科技纵横(2021年24期)2021-03-02
疯狂英语·新悦读(2020年7期)2020-07-30
癌变·畸变·突变(2020年1期)2020-02-12
中成药(2018年6期)2018-07-11
山西医科大学学报(2017年11期)2017-12-01
天津科技大学学报(2016年3期)2016-08-02
食品研究与开发(2016年9期)2016-06-13
哈尔滨商业大学学报(自然科学版)(2016年1期)2016-04-22
上海蔬菜(2015年2期)2015-12-26
