
纳米抗体的稳定性及其结构基础研究进展*
2022-07-21 11:51何晓婷董洁娴沈玉栋徐振林
生物化学与生物物理进展 2022年6期
何晓婷 董洁娴 沈 兴 王 弘 沈玉栋 徐振林**
(1)华南农业大学食品学院,广东省食品质量安全重点实验室,广州 510642;2)深圳清华大学研究院抗肿瘤创新药物研发中心,深圳 518057)
驼类动物体内含有3种不同亚型的抗体,分别是常规抗体IgG1、天然缺失轻链及CH1恒定域的重链 抗 体(heavy‑chain antibody,HCAb) IgG2与IgG3。克隆并表达重链抗体可变区可得到一个晶体结构为直径2.5 nm、长4 nm 的椭球形单域抗体(图1),其分子质量小到只有免疫球蛋白的1/10(~15 ku)却保留了全部的抗原结合能力,因此也被称为纳米抗体(nanobodies,Nbs)或VHH 抗体(variable domain of the heavy chain of heavy‑chain antibody)[1]。与传统抗体片段如抗原结合片段(fragment of antigen binding,Fab) 和单链抗体(single chain antibody fragment,scFv)相比,Nbs具有多个明显的优势,比如免疫原性弱、生产成本低、水溶性好、组织渗透性好、稳定性与亲和力较高等[2]。正是这些优良特性使得Nbs在生物技术方面得到了非常广泛的应用[3‑6]。
对于大多数应用而言,稳定性是制约抗体应用的关键因素之一。一方面抗体在其生产、运输、储存及使用的过程中容易发生多种物理和化学降解,另一方面抗体的不稳定性聚集会潜在地影响产量、保质期和免疫原性等应用参数[7]。因此抗体的稳定性不仅会影响其生物学和生化评估,还会影响纯化、储存以及配方设计和生产。在各种类型的抗体中,Nbs显示出极好的溶解性,并且对高温和化学变性具有显著的抗性,可以很大程度地克服传统抗体片段scFv 的聚集和降解等稳定性问题,是许多应用的理想选择。
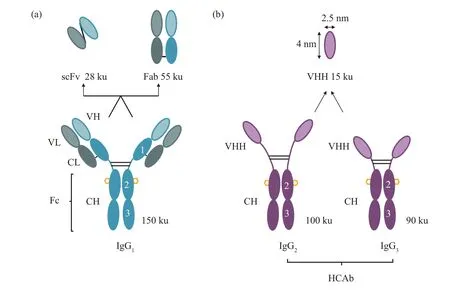
Fig.1 Structures and molecular mass of three different subtypes of antibodies and antibody fragments in camelid animals图1 骆驼科动物体内3种不同亚型抗体与其抗体片段的结构与分子质量大小
虽然Nbs与传统抗体相比在稳定性方面普遍具有明显的优势,但是不同的Nbs 稳定性差异较大,主要与其特殊结构有关。已经有很多研究者基于共有序列的分析基础上对Nbs进行改造来进一步提高稳定性,同时也鉴定出降低Nbs聚集的共有结构特征[8‑11]。本文在前人对Nbs的稳定性表征以及结构分析的基础上对其在高温、有机溶剂以及其他极端条件下的稳定性表现进行了系统性总结,并结合稳定性数据和序列信息揭示了高度稳定的Nbs具有较高的净电荷表面、限制构象迁移的二硫键、亲水氨基酸取代较多的框架区、保守的疏水性口袋以及高度相互作用的结构域等结构特征。基于这些结构特征,本文还讨论了几种Nbs 的稳定性结构优化策略,包括共有序列驱动的序列修复、替换易于修饰的氨基酸、净蛋白质电荷的改变、非天然二硫键的引入和互补决定区(complementarity determining region,CDR)的移植。值得注意的是,这些策略不是绝对通用的,有可能以牺牲亲和力或者产量为代价,因此在进行结构稳定性优化时要综合考虑稳定性、亲和力以及表达量等多方面的因素。
1 纳米抗体结构
Nbs 与常规抗体可变区(variable region of heavy chain,VH)的空间结构相似,其框架是由9个反向平行的β 折叠片层(A‑B‑C‑D‑E‑F‑G‑H‑I)通过链间氢键和二硫键连接在一起组成(图2a)[12]。在此结构中,3 个CDR 分别连接BC、DE和HI 链,并靠N 端形成连续表面,与抗原表位的表面互补[13]。连接CDR之间的氨基酸序列相对比较保守,称为骨架区(framework region,FR)。几乎所有Nbs 结构都含有一个连接FR1(C23)和FR3(C104)的保守二硫键,该键跨越蛋白质的内部,将两个β链连接起来,增加了结构稳定性[11]。部分Nbs 还含有一个可限制CDR 环柔韧性和构象自由度的额外二硫键[14]。
传统抗体VH 和VL 通过疏水作用力来稳定结构并共同构成抗原结合区。与之相比,Nbs仅含有3 个可变区,尽管与抗原结合界面的表面积减少,但其仍具有较高的稳定性与亲和力。它主要通过以下方式来适应轻链的缺失:a.大量的亲水性氨基酸取代先前与CH1、VL 结合界面的脂肪族残基(L12S、V42F/Y、G49E、L50R/C、W52G/L),且部分FR2被拉伸扭转的CDR3环覆盖,避免与外界水环境的接触[12,15],从而防止Nbs 的二聚化,其CDR3 环 越 长,Nbs 越 稳 定[7,16‑17]。另 外 还 可 在CDR3 末端形成疏水核心,有利于稳定Nbs 的折叠结构域[18]。b.Nbs 的CDR1 和CDR3 普遍比VH 的长(图2b),潜在地增加了互补位构象的多样性,从而以高度的形状表面互补性与相应的抗原结合,一定程度上弥补了轻链缺失造成的抗原结合力下降以及因尺寸小而导致的潜在序列多样性降低[19]。
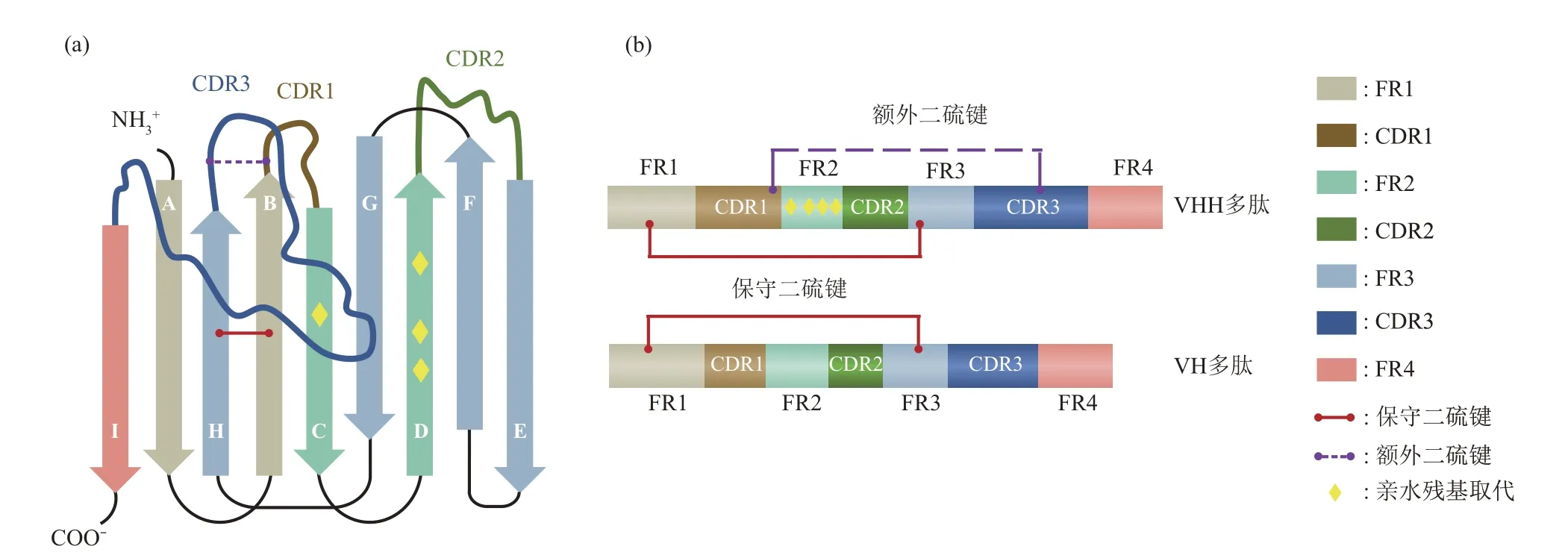
Fig.2 Structure of Nbs图2 Nbs的结构
2 稳定性表征
抗体的物理和化学稳定性的全面表征对于设计稳定的抗体制剂以实现所需的功效和保质期至关重要。在抗体的研究、生产与应用中,诱导变性因素有很多,包括温度、有机溶剂、压力、化学试剂、酸碱环境以及蛋白酶等,可以基于这些因素采用互补或独立的方法来表征Nbs的稳定性。
2.1 热稳定性
Nbs的一个固有特性是其热稳定性,包括出色的热展开抗性以及变性后重新折叠的能力。部分Nbs可以在-20℃、4℃条件下保存几个月,甚至在37℃条件下温育几个月到一年也不会丢失其抗原结合特性[8,20],这种长期稳定性使其能在室温下运输、储存和使用。当温度逐渐升高时,抗体会趋于热变性展开,一般使用解链温度(Tm)推断抗体的展开倾向。Nbs 的Tm覆盖了50~80°C 的广泛范围,普遍显示出对热诱导具有较高的抵抗力[21]。抗葡萄球菌肠毒素B的Nb A3是一个比较经典的例子,其Tm高达85℃,是迄今为止报道的最高纪录[22]。这种热展开抗性使得Nbs 可以应用于相对较高的温度中,比如在50℃条件下对Nb进行放射性标记,以实现对动脉粥样硬化炎症标志物的监测[23]。
Fab和scFv等常规抗体片段的Tm测得值范围基本与Nbs相似,说明Nbs和常规抗体片段可能显示出相当的热平衡稳定性[24],但是使Nbs 区别于常规抗体的主要标志是其在极端温度下的可逆性[25‑26]。这种可逆性可以通过检测高温孵育一段时间后Nbs 的活性来进行表征。对部分已报道的Nbs 免疫分析文献进行总结,发现大部分Nbs 在85℃高温长时间处理后仍可保持结合活性,而其他常规抗体则不可逆地失活[27‑38](表1)。例如,抗咖啡因Nb可在90°C的温度孵育后保留90%的活性,但其单克隆抗体(monoclonal antibody,mAb)在70°C 孵育后会失活[28]。这种耐高温咖啡因Nb在一次性侧向流动装置的应用中已获得专利,可用于监测热饮料中的咖啡因[39]。同样地,针对黄曲霉毒素的Nb 以及抗独特型抗体(anti‑idiotype antibody,Aid)耐热性也优于常规抗体,在高温条件下短时间孵育后其多克隆抗体(polyclonal antibody,pAb)与单克隆抗体的结合活性仅保留20%甚至完全丧失,但在相同温度条件下孵育20 min甚至1 h后Nbs仍具有高达40%~80%的结合活性[34,36,38],这使其可用于涉及高温过程的黄曲霉毒素检测,比如需要巴氏灭菌的乳制品等。总而言之,这些特性使Nbs在热变性的复杂条件下比传统抗体具有更广泛的应用优势。
2.2 有机溶剂耐受性
在开发用于亲脂分析物的免疫分析方法时,一般采用有机溶剂对分析物进行充分提取,但是有机溶剂会影响抗体的活性,因而在前处理步骤中需要进行蒸发复溶或者高倍比稀释来避免有机溶剂的影响。优选有机溶剂耐受性较高的抗体可以省去蒸发复溶的步骤或避免高倍数稀释导致分析灵敏度的下降。对部分Nbs的有机溶剂耐受性评估进行总结发现大部分Nbs 可耐受较高浓度的甲醇、乙醇、乙腈、丙酮或二甲基亚砜[27,34,37,40‑43](表2),而这些溶剂是提取高度亲脂性分析物的首选溶剂。例如,从食品基质中提取赭曲霉毒素A(OTA)需要用到甲醇,但是甲醇不仅会影响了OTA 与抗体的相互作用,还会抑制免疫测定中的二抗活性[44‑45]。在20%的甲醇溶液中,抗OTA的Nb在测定时仅稀释2.5倍即可消除样品基质干扰,但是其mAb测定需要稀释30 倍以上才能消除,表明了Nbs 比传统抗体对样品基质干扰有更高的抵抗力[37]。当高度亲脂性的黄曲霉毒素作为目标分析物时,Nb 能在80%的甲醇溶液中保持100%的结合能力,而其mAb 在相同条件下则会失去50%的活性[34]。这种耐有机溶剂特性不仅有利于检测需要从样品中萃取的亲脂性分析物,还可以使其应用于需要较少稀释的亲和柱开发[34]。

Table 1 Comparison of heat resistance between Nbs and conventional antibody表1 Nbs与常规抗体耐热性的比较
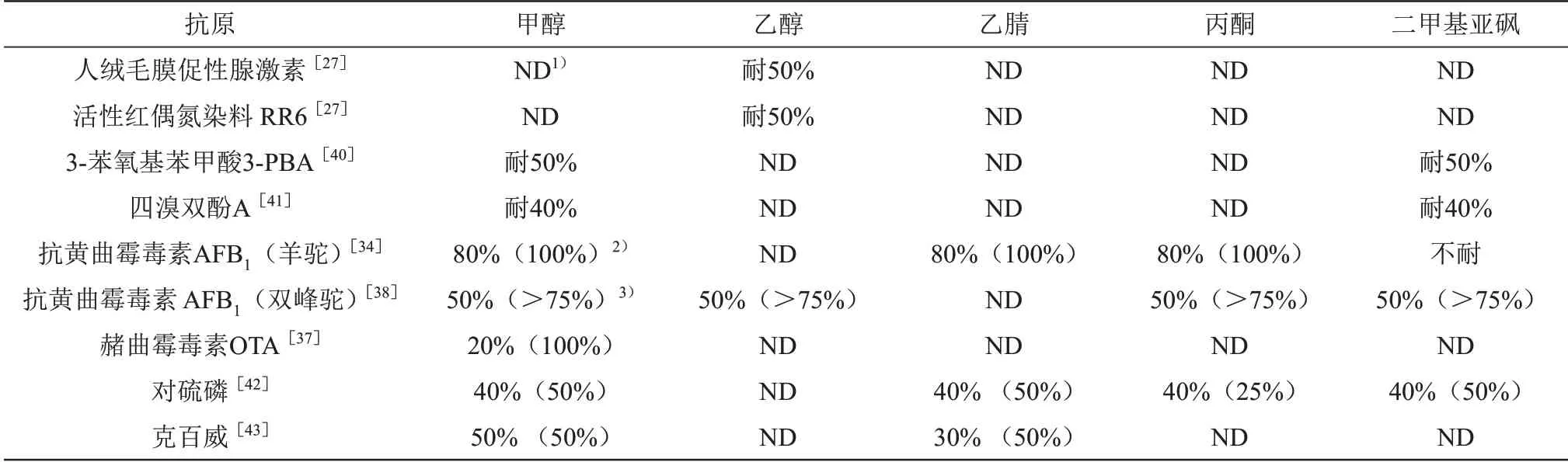
Table 2 Tolerance of Nbs in different organic solutions表2 Nbs在不同有机溶液中的耐受性
2.3 压力耐受性
在压力耐受性方面,Dumoulin等[24]报告了针对人溶菌酶与偶氮染料RR6的Nbs在大于400 MPa的压力条件下才会使其变性展开,说明了Nb 在极端压力条件下的稳定性。另外,在较低的压力条件(约50~250 MPa)下可以加速解离非共价蛋白化合物,因此可在这个压力范围内分离抗原‑Nbs 复合物而不会破坏抗体。该策略可用于免疫亲和分离,在受控的压力增加后从免疫吸附剂中洗脱特异性结合的抗体。此外,这种可耐受极端压力以及小体积和高溶解的特性还有利于Nbs开发为雾化吸入制剂用于呼吸系统疾病的治疗。比如现阶段已经进入临床II期研究的三聚体Nb ALX‑0171可通过气溶胶递送方式有效降低住院婴儿的呼吸道合胞病毒载量[46]。同为三聚体Nb的PiN‑21也成功通过喷雾给药的方式以超低剂量预防并治疗了仓鼠动物模型中的SARS‑CoV‑2冠状病毒[47]。
2.4 化学试剂耐受性
一般化学变性剂有盐酸弧、尿素、硫氰酸铵溶液等。除了压力稳定性测试,Dumoulin等[24]还探究了Nbs的化学诱导展开实验,实验表明Nbs的盐酸弧、尿素变性浓度分别在2.3~3.3 mol/L 和高于6 mol/L 的范围,明显高于常规抗体scFv 片段(1~2 mol/L和2~3 mol/L)。与之类似,针对hCG与RR6 的Nbs 所需的变性硫氰酸铵浓度也比其mAb高,其中最稳定的Nb在4 mol/L硫氰酸铵溶液中仅损失20%的生物活性[27,48]。另外,在洗发剂存在条件下分离出的Nb 不仅可以在含有高浓度表面活性剂的洗发水中稳定结合抗原马拉色菌,在高浓度的尿素与盐酸胍条件下也保持稳定,可将该Nb 与洗发剂混合使用来防止头皮屑[48]。但是并非所有化学变性剂对抗体反应的影响都是消极的,比如抗李斯特菌Nb 在高尿素浓度下会提高抗原结合力,表明尿素有助于部分Nbs的重新折叠[49]。
2.5 极端pH耐受性
Nbs在不同pH值条件下也具有相似的稳定性。比如,Wang 等[41]报道的抗四溴双酚A 的Nb 抗体在pH 值为7.4~10 之间非常稳定。同样地,针对李斯特菌的Nb在pH为2及12的酸碱极端条件下孵育过夜后仍能保留至少30%的活性[49]。另外,酸碱性条件的变化对抗皮质醇Nb 的抗原结合力也不会有明显的影响,使得该Nb 抗体在微流体实验中具有受测试条件可变性影响较小的优势[9]。
2.6 蛋白酶抗性
部分Nbs还具有较强的蛋白酶抗性,可抵抗胃肠道中的蛋白酶解,适于用作口服给药的治疗剂。通过DNA改组和随机诱变得到的稳定性Nb可在肠液或胃液培养后仍分别保持90%和41%的活性[50]。在蛋白酶压力下淘选出来的Nb 也具有优秀的蛋白酶抗性,能显著减少鸡胃肠道中的空肠弯曲杆菌[51],可作为一种替代抗生素的潜在饲料添加剂加以推广应用。此外,这种可规避蛋白酶解的特性还有利于开发靶向细胞内抗原的功能性重组抗体Nbs,使其成为对抗细胞内病原体或治疗神经退行性疾病的新型治疗剂[52]。
3 稳定性的结构基础
Nbs的小分子和单域属性赋予了其比常规抗体更为稳定的特性,然而并非所有的Nbs都具有很好的热稳定性与化学稳定性。在以往研究的多个Nbs中,大约有2/3 的Nbs 在65℃处理后发生了不可逆的聚集,但是部分Nbs 在90℃高温处理后仍具有90%的活性[7]。这些稳定性差异的根本原因在于Nbs特殊结构的差异,包括氨基酸序列、二硫键的数量与位置、结构域的构象等。
3.1 氨基酸
3.1.1 氨基酸的化学修饰
尽管Nbs可以通过原核生物进行大量表达,没有翻译后修饰的过程,但是在抗体储存、生产与应用过程中的一些胁迫条件容易使抗体的一些氨基酸发生化学修饰。其中天冬氨酸(Asn)与谷氨酰胺(Gln)的脱酰胺反应是较为重要的一种化学修饰,以Asn的脱酰胺更为普遍。这些化学修饰的积累会改变抗体的局部疏水性以及空间结构从而影响其生物活性,其中抗体CDR 区的脱酰胺作用还可能会导致抗原结合功能的丧失。Akazawa‑Ogawa 等[8]还通过实验验证了化学修饰是导致Nbs热诱导不可逆变性的主要驱动力,其Asn残基的化学修饰会改变抗体的范德华相互作用和天然结构的氢键,从而导致Nbs的稳定性和折叠能力发生变化。除了Asn修饰,热诱导还会引起半胱氨酸(Cys)的化学修饰与二硫键的交换。研究表明对野生型抗hCG Nb进行热处理,Cys的修饰会导致巯基数量的减少以及Cys残基周围的断裂,这可能是具有单个二硫键的Nbs 变性的原因[53]。这些易于发生化学修饰的氨基酸是影响Nbs稳定性的关键氨基酸,可能会引起抗体变性后折叠能力的逐渐丧失。
3.1.2 氨基酸的电荷
增加多肽链的净电荷会增强溶解度并导致未折叠状态的分子间排斥,提高抗体对聚集的抵抗力[54]。与VH 抗体相比,D62、K65、R67、R72、K76和E89在Nbs结构域中出现得更频繁,这些带电荷残基的普遍使用增加了电荷排斥,降低了Nbs聚集的倾向,使得Nbs蛋白可逆折叠、热稳定性和可溶性的性质优于传统抗体[17,19]。据报道[55],具有抗聚集特性的VHs 与Nbs 的等电点pI 多倾向于酸性,这表明含有大量负电荷残基可能是Nbs稳定性和溶解性进化的内在因素。此外,Nbs序列中一些保守性的带正电荷残基可增强与介质中存在的带负电荷分子的静电相互作用,比如皮质醇分子被其Nb中几个带正电荷的残基紧密包围,增加了Nb在恶劣环境中与抗原的结合稳定性[56]。又比如与糠秕马拉色菌结合的Nb 序列中带正电荷的残基(R/K44)对于Nb 在高pH、洗发剂、尿素和盐酸胍等条件下与真菌的结合至关重要[48]。以上表明具有高稳定性的Nbs趋于含有更多净电荷,尤其含有更多带负电荷的氨基酸,而结构中带正电的残基可通过静电作用来促进抗原与抗体之间的稳定性结合。
3.2 二硫键
二硫键在分泌蛋白的折叠和稳定性中起着关键作用。据报道[9,57],使用大肠杆菌系统表达Nbs时,在还原性细胞质中产生Nbs 的Tm要比在周质的氧化环境中产生的Nbs低得多,这种稳定性差异主要与二硫键的形成有关[21,58]。在氧化环境中,Cys残基的成对氧化将二硫键引入多肽链中,通过约束蛋白质的未折叠构象来提高折叠结构域的机械稳定性及构象稳定性[53,59‑60]。在Nbs结构中,大多会出现1个及以上的二硫键,包括连接FR1(C23)和FR3(C104)的保守二硫键以及连接CDR3与其他结构域的额外二硫键,这些二硫键是固定Nbs三级结构的关键结构之一。
3.2.1 保守二硫键
Nbs的稳定性高度依赖于这个几乎所有Nbs结构共有的保守二硫键。该域内二硫键埋藏在Nbs折叠域的疏水核内部,将两个β折叠连接在一起,使Nbs 肽链的空间结构更为紧密[60]。通过定点诱变除去野生型Nbs的天然保守二硫键可以显着降低其Tm(15~25°C)[9,53,59,61],表明了该键是影响Nbs平衡热力学稳定性的关键结构。但是该键不是Nbs结构与功能所必需的,如果Nbs具有较强的固有稳定性或已通过蛋白质工程进行稳定性改造,则该键的缺失与替换不会影响其结构稳定性与抗原结合能力[60]。比如衍生自单峰骆驼的通用Nbs 框架cAbBCII10在去除保守的二硫键后仍具有良好的表达水平与稳定性[62]。此外,还有文献报道了部分无二硫键的Nbs突变体可以折叠成原生结构,保留其抗原结合能力,也不会影响其抗体抗原结合复合物的刚性[53,59,61,63]。
3.2.2 额外二硫键
除了保守二硫键,部分Nbs还含有一个可以稳定可变区空间构象的额外二硫键[64]。在研究中发现,在两种同源Nbs中,含有双二硫键的Nbs要比仅含有保守二硫键的Nbs具有更高的热稳定性与构象稳定性[59]。同样地,对大量的Nbs 序列对比分析发现具有额外二硫键的Nbs 的Tm值平均会高5.2℃[65]。这些额外的二硫键约束并稳定增大的CDR3环,限制构象迁移并防止其热诱导聚集,提高了其构象稳定性[11‑12,14,66]。此外,额外二硫键会通过降低Nbs的聚集速率来减少不可逆聚集,提高了动力学稳定性[20]。主流观点认为这个额外二硫键还可以通过降低其与抗原结合的熵损失来提高结合亲和力[14]。但是,Mendoza等[17]对此进行了反驳,他们分离出一种对亲和力没有明显变化的二硫键突变体。由此可见,这个额外的二硫键可能已经进化成为稳定Nbs结构域的生物物理特性,在特定情况下可在抗原结合中发挥一定的作用,但是这种作用不具有广泛的普遍性。
根据编码Cys的位置可以将其额外二硫键大致分为3 种不同类型(图3),分别是连接CDR3 与CDR1 或CDR2/FR2 边界的环间二硫键以及CDR3的环内二硫键[12]。不同驼类在额外二硫键的位置与丰度上有很大的差异。据报道[17],骆驼中额外二硫键的出现频率约30%,比羊驼(~12%)和美洲驼(~3.5%)高。这是因为大部分羊驼Nbs 因其CDR3 环相对较短而不存在这种额外的二硫键[67],只有小部分羊驼Nbs 在FR2(C55)和CDR3 间存在额外增多的Cys[66,68]。另外,美洲驼抗体仅一个特殊亚科VHH3 含有延伸的CDR3,该CDR3 被一个位于FR2(C50)的二硫键连接,但是这个亚科的Nbs很少被分离出来,因此文献中报道的绝大多数来自美洲驼的Nbs 仅在保守位置出现二硫键[11,69]。而骆驼Nbs 中延伸的CDR3 则一般是被位于CDR1(C31/33/34)的二硫键约束[11,70],其另一个Cys 出现在位置FR2(C50)或CDR3 上的Nbs 也曾见于报道[67,71]。这说明了这个额外的二硫键不是维持其稳定性的必要结构,具有不同生态适应性的驼种Nbs根据其序列及其长度的差异采用不同数量以及不同位置的二硫键来丰富其稳定性机制。
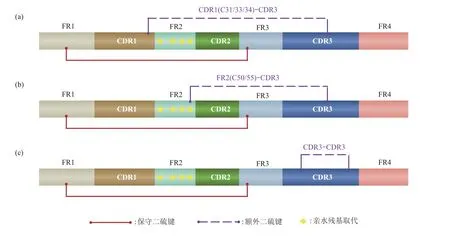
Fig.3 Three different types of additional disulfide bonds in Nbs图3 Nbs中三种不同类型的额外二硫键
根据以上分析可以看出在CDR1、FR2、CDR2区域出现的Cys 一般出现在保守位置上,而CDR3的Cys位置是随机的,这是因为前者是由骆驼的种系基因编码,而CDR3中第二个Cys密码子出现在V‑D‑J重排后或之后的体细胞超突变[72]。大量序列分析发现,Nbs的Cys总是成对出现的,这是由于Nbs 在B 细胞受体成熟期间进行阳性选择的结果[14,59,73]。含有未配对Cys 的Nbs 容易自发氧化形成二聚体而失去抗原结合能力,即便这种克隆被成功分离出来也会因为长时间储存而导致二聚化[74]。但是,存在一种特殊情况允许这种单Cys的存在,最近报告了一种含有金属离子结合位点的Nb,该Nb 结构中没有配对的Cys 与其他3 个残基形成了一个与金属离子Zn2+配位的几何结构,其中Zn2+代替二硫键充当CDR1 和CDR3 环之间的桥,为CDR3环构象提供了更大的刚性[72]。
3.3 结构域
3.3.1 单体结构
对于Nbs来说,其小分子和单域属性是其耐受温度的主要机制。已经证明了同源的常规IgG1抗体、HCAb、Nbs片段热稳定性依次递增[26,75],这可能由于Nbs只有1个免疫球蛋白折叠域,没有轻链、CH1、铰链区以及Fc片段,其分子质量与结构复杂性要比分别具有6与12个免疫球蛋白折叠域的HcAb、IgG1抗体要低。在应对高温等变性条件时,理论上具有较少折叠形式的蛋白质片段更容易重新折叠从而具有更高的耐受性[76]。此外,常规抗体的二级结构很容易形成增加聚集倾向的β 折叠结构,且其重轻链疏水结合界面的暴露在不可逆聚集中起着核心作用,这些结构特征都导致了骆驼血清中的重轻链抗体IgG1在热处理后其结合活性严重损失[26,77]。在构建scFv文库时,VH与VL的结合容易出现错配,导致其在重组表达系统中表达不佳,出现无法识别抗原以及结构不稳定等问题[20]。相比之下,只含1个结构域的Nbs不受重链与轻链配对的限制,避免了这种结构解离导致的不可逆聚集以及重组表达的错配,大大提高其在变性后或者表达的折叠效率,从而提高了理化稳定性。
3.3.2 FR框架区
CDR 主要决定了Nbs 对靶标的特异性和亲和力,但是针对化学变性剂、热、蛋白酶和极端pH的稳定性主要取决于结构域中保守的FR 序列。在传统抗体中,FR 区含有大量的疏水残基(L12、V42、G49、L50、W52、Y95和W117),介导了重链与CH1、轻链的结合[19,67,78]。当其孵育温度高于Tm时,这个疏水界面会随着常规抗体天然结构的解离而暴露在外,可能引起分子间的相互作用,导致不良的非特异性聚集与沉淀[24]。Nbs 的聚集需要蛋白质展开,从而强调了天然Nbs 的高溶解度[7]。在骆驼科动物Nbs中,为防止重链聚集,框架区内的一些残基突变为更亲水的残基(L12S、G49E、L50R和W117R/G),而其他突变(V42F和W52F/G)则被较长的CDR3 环覆盖从而屏蔽了溶剂[19,67,78‑79]。特别地,V42F 突变可以填充VH 中由Y95、W117 和CDR3 的侧链形成的疏水口袋,而Nbs中的W117R取代允许F42较大的侧链向Y95和R117转移[80],增强结构内部的疏水堆积,从而增强其结构稳定性[81]。除此,FR1 的5V 和6E 残基是很多稳定Nbs的共有氨基酸序列,已经成为引入负突变工程化的通用残基[82]。此外,对人VH片段的氨基酸进行“骆驼化”突变,其熔点高于野生VH片段的实验也证明了Nbs框架序列中的特定氨基酸有助于提高温度稳定性[83]。以上研究表明,特定的氨基酸取代和重新定向的侧链可以重塑Nbs结构域并增加其亲水性。
3.3.3 CDR超变区
Nbs与常规抗体的另一个区别特征是明显延伸的CDR3 环。序列分析验证了Nbs 的CDR3 平均长度比常规抗体多5 个氨基酸[84],尤其是单峰骆驼具有更长的CDR3环[85]。这些长CDR3环可以增加抗体与抗原的接触面积,在一定程度上弥补轻链缺失造成的抗原结合力下降的不足[69]。理论上,这个增大的CDR3环表明其具有更大的柔韧性,这可能会降低结合亲和力和抗体稳定性。但是在实际的研究中发现,CDR3长度和热稳定性之间并没有预期的负相关性,而是随着CDR3长度的增加,稳定性略有增加的趋势更加明显[7]。较为合理的解释是长的CDR3环更有利于通过与残基的相互作用屏蔽VH 与VL 结合的疏水界面,增加其亲水性并防止二聚化。同时,这种非共价相互作用也稳定了这个长CDR3环,补偿了长环CDR3区可能引起的不稳定效应。
此外,CDR3 环N/C 端附近的Y93、F/Y117 、W118 等保守氨基酸形成的疏水簇也有助于稳定CDR3 环的构象[18,67]。因此,CDR 区残基可以与FR 区残基相互作用,并且这种相互作用具有中和潜在聚集点和对互补位施加结构刚性的双重作用,其稳定性与CDR3长度正相关,通过二硫键或者其他非共价相互作用稳定长CDR3环可进一步增强其稳定效应。
4 结构的稳定性优化
Nbs稳定性信息与结构信息的合并揭示了高度稳定纳米抗体所特有的结构特征。基于这些结构特征可以对Nbs采用合理的设计方法来进行结构的稳定性优化,包括共有序列驱动的序列修复、替换易于修饰的氨基酸、净蛋白质电荷的改变、非天然二硫键的引入和CDR 的移植。通过这些策略开发出在极端条件下仍能保持结合能力的稳定Nbs,从而为生物技术、检测、诊断和治疗应用提供高性能的试剂。
4.1 点突变
4.1.1 基于共有序列的诱变
同一个对象可以筛出多株序列不同的Nbs,这些序列的FR 区的某些位置基本保守不变,如果这些保守残基发生变化可能会导致同源Nbs 的Tm出现重大差异[86‑87],当恢复这些保守序列有可能导致Tm的增加。例如,一株抗蓖麻毒蛋白Nb 在第117位残基发生偏差从共有序列Trp变为Arg,恢复此突变可使其Tm增加而不会降低其抗原亲和力以及产量[10]。但是该方法也有例外,比如有研究者通过I122T 突变来恢复抗肉毒杆菌神经毒素Nb E7在位置122的共有序列IIe,虽然可以提高产量,但是突变体Tm并没有发生变化[76]。虽然该策略将稳定的高概率归因于蛋白质家族特定序列位置上最常见的残基,具有一定的物种依赖性与偶然性,但是可以作为一个稳定性优化的重要方向。
4.1.2 易修饰残基的替换
对抗体序列中易受化学修饰的位点进行结构改造也是其稳定性优化的重要方向之一。据报道[8],用其他的氨基酸取代Nb 序列中所有易于修饰的Asn 残基(N57S/N82S/N92T)所得到的突变体比野生型Nb 更能抵抗热诱导的不可逆变性。此外,替换未配对的Cys不仅可以提高产量,还可能提高其Tm[10]。Asn和Cys的修饰是引起Nbs变性的关键因素之一,替换这些易于修饰的残基有利于提高Nbs的耐热性。
4.1.3 负突变残基的引入
通过用带电残基替换暴露于溶剂的残基来增加蛋白质的净电荷,可以显着降低其聚集特性。由于稳定性Nbs的等电点倾向于酸性,因此在某些Nbs的序列中引入负电荷突变或者在其C端引入一段负电荷序列以降低等电点,不仅可以提高产量,还可以提高溶解度、熔解温度与抗聚集能力,改善变性后的折叠,利于Nbs 在高浓度条件下的储存[10,65,76,82]。添加电荷可以防止聚集,但也会破坏抗体的折叠状态,选择要突变的残基要同时保留Nbs 天然结构,在非抗原结合区的框架1 中引入负突变Q1E/D、K3Q、Q5V、A6E基本成为了优先考虑的负突变位点[10,65,82,88]。
4.2 二硫键的工程化
在Nbs中进行二硫键的工程化是提高其稳定性最为广泛使用的方法之一,大量文献集中研究比较了引入不同数量与位置的Cys对Nbs的稳定性和亲和力的影响(表3)。在数量上,Saerens等[59]分别在3 个不同的Nbs 中通过突变获得了含有0~3 个二硫键的突变体。实验表明,去除保守二硫键大多会导致其Tm的急剧下降,而引入一个工程化二硫键则基本可以提高其稳定性,但是当Cys 数量达到8个甚至10 个时则容易导致突变体形成无功能多聚体。此外,目前也鲜见具有多于4 个天然Cys 的Nbs 的报道,说明过多Cys 容易导致二硫键的异常配对,无法形成正确稳定的三级结构。在位置上,Hagihara等[9]首次使用Cys取代Nbs中的埋藏残基A54(FR2)和I78(FR3),插入的二硫键在抗原亲和力基本不变的情况下使其Tm提高了10°C。其中残基A54 和I78 在Nbs 中高度保守,并跨越β 折叠之间的疏水内部。随后有不少研究人员相继对不同抗体的相同位置进行了突变,加入第2个二硫键的突变体可使其Tm增加4~20°C,甚至在多次加热冷却循环处理后仍保持接近100% 的结合能力[10‑11,89]。此外,引入这个连接FR2与FR3的二硫键还可以在酸性pH 和存在化学变性剂的情况下提高Nbs热稳定性和蛋白酶抗性,使其具有更高的胃肠道稳定性[51,89‑91]。另外,也有学者比较了其他不同二硫键插入位点(C4~117、C36~110、C53~112以及C44~50)对其稳定性的影响,但是发现尽管大部分突变体都可以提高亲本的Tm,但却牺牲了部分亲和力、产量或者变性重折叠能力[11,92]。以上实验表明通过蛋白质工程在A54 和I78 处引入额外的二硫键已经成为提高Nbs平衡热力学稳定性的通用方法。
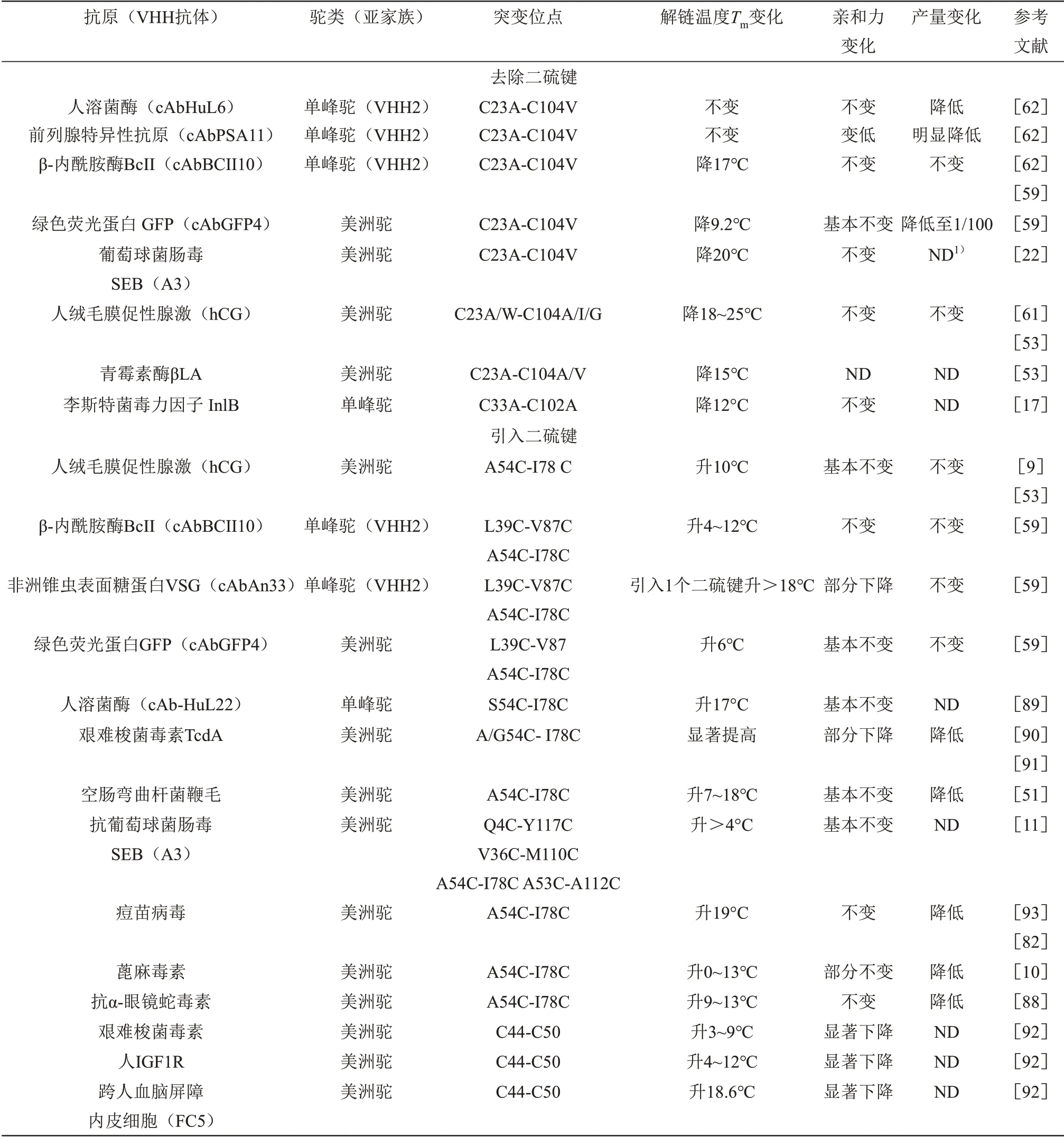
Table 3 Disulfide bond engineering of Nbs表3 Nbs二硫键的工程化
通过定点诱变加入工程化的二硫键是稳定Nbs的有效方法,但是此方法也会对其应用性能产生一定的影响。a.在需要高温长时间孵育的应用中,添加的额外二硫键甚至天然存在的二硫键可能会由于热诱导的二硫键改组和Cys残基的修饰,显示出影响Nbs复性的能力[20‑21,53]。b.在某些情况下,二硫键的引入会对亲和力以及特异性产生难以预测的负面影响[59,90]。c.由于在折叠过程中可能形成错误的二硫键,Nbs在大肠杆菌中的表达会受到二硫键的影响[14,21]。因此,在进行抗体的二硫键工程化时应该综合考虑稳定性、亲和力与表达量等方面的影响。
4.3 CDR超变区的移植
由于部分Nbs序列高度保守,可以通过移植不太稳定的抗原特异性Nb的CDR到一些高度稳定的Nbs 框架上以获得具有环供体Nb 的抗原结合特性与支架受体Nb的稳定性的嵌合体,这种CDR移植技术已用于常规抗体片段的稳定改造[94‑95]。在Nbs的应用中,已经报道了多个可以实现相应功能的保守框架,例如人源化框架cAbBCII10[96]、hs2dAb[97]、C8WT[98]以及针对细菌表达优化的人工设计CF支架[99]。其中框架cAbBCII10是一种具有较高构象稳定性的单峰驼通用框架,将家族2 Nbs 的CDR 移植到该框架上保留了CDR 供体抗原结合能力的同时显示出了增加的稳定性[62]。此外,Zabetakis 等[100]此前曾报道,将CDR2 从具有高熔点的抗葡萄球菌肠毒素B的Nb A3移植到具有低熔点的抗蓖麻蛋白Nb中可以使嵌合体克隆Tm提升15℃。虽然CDR 移植比较局限于Nbs 亚家族的限制[62],且还可能导致Nbs 与靶标结合能力的下降[100],但是利用具有特定特性的支架进行CDR超变区交换是一种尝试增加抗体稳定性的可行方法。
5 展 望
虽然目前已经有很多研究者通过定点突变、引入二硫键等工程化的方法制备出很多相对亲本更稳定的Nbs,但是大多数的工程化是基于共有序列分析与经验值的基础上,鲜有报道涉及晶体结构的稳定性分析,这可能导致得出的稳定性结构信息具有一定的片面性。蛋白质晶体的制备与X‑射线衍射晶体结构解析是研究蛋白质结构最直观、准确的技术,可以在分子水平上阐明分子间相互作用的结构基础。未来的研究可以更多地综合利用蛋白质结晶、X‑射线晶体结构解析、分子动力学等技术建立Nbs与抗原的三维空间结构模型,明确重链抗体识别方式、关键氨基酸残基、结合位点、作用力类型等关键信息,全面探讨Nbs的一级序列结构、二级折叠结构、三级空间结构与稳定性的关系与规律。鉴于Nbs在临床应用上取得的成果,未来Nbs在复杂基质与环境的检测等方面具有较为广阔的应用前景,值得进行深入的系统研究。
猜你喜欢
建材发展导向(2022年20期)2022-11-03
中国临床新医学(2022年8期)2022-09-08
保健与生活(2022年12期)2022-06-09
文萃报·周五版(2022年12期)2022-04-02
健康之家(2021年2期)2021-07-01
华东师范大学学报(自然科学版)(2019年5期)2019-11-11
祝您健康(2018年12期)2018-11-27
新农业(2018年3期)2018-07-08
电机与控制学报(2018年9期)2018-05-14
家庭用药(2018年12期)2018-02-14
