
益生菌中胆盐水解酶作用机理研究现状
2010-01-04 11:34任婧吴正钧王荫榆
中国乳品工业 2010年2期
任婧,吴正钧,王荫榆
(光明乳业股份有限公司技术中心基础研究部,上海 200436)
益生菌中胆盐水解酶作用机理研究现状
任婧,吴正钧,王荫榆
(光明乳业股份有限公司技术中心基础研究部,上海 200436)
对乳酸菌中胆盐水解酶研究现状进行分析,从编码基因、蛋白氨基酸结构以及酶与肠道间密切关系等方面入手,阐述了肠道益生菌通过胆盐水解酶对胆盐的水解,可以达到耐受肠道胆盐压力,增强细菌表面对体内免疫系统的抵抗能力建立长期感染,促进在高等动物肠道内细菌的定殖,降低血清胆固醇水平等作用,为最终治愈高胆固醇血症提供了良好的解决方案。
胆盐;胆盐水解酶;bsh益生菌;高胆固醇血症
0 引 言
益生菌是指那些对人、动物产生有益影响的活性微生物,通过摄入一定的数量后,对宿主可产生一种或多种特殊的、超越其固有基本营养价值的保健作用。动物对益生菌具有选择性,通常体现在很多物理压力方面,如低pH值、耐胆汁、以及渗透压和功能性选择等;而可以耐受胆盐是益生菌被选择的重要特征。本文综述了益生菌中胆盐水解酶作用机理研究现状。
1 胆酸循环及胆盐水解过程
在哺乳动物中胆固醇(cholesterol)代谢终产物就是在肝脏中从头合成胆汁酸。胆汁酸通过与牛磺酸(taurine)或甘氨酸(glycine)交联形成胆盐(bile salt),然后随胆汁一起分泌。胆汁是一种黄绿色溶液,主要成分除胆汁酸外还包括胆固醇、磷脂以及胆绿素等。它的功能主要是作为生物溶剂乳化和溶解脂肪,因此在脂肪代谢中具有重要意义。另外它也具有潜在的抗菌活性,因为可以溶解微生物膜。初级胆汁酸,包括胆酸(cholic acid)及牛黄鹅去氧胆酸(chenodeoxycholicacid)在肝脏中从胆固醇从头合成。疏水的甾醇核通过与甘氨酸或牛磺酸交联可溶性增加,因此形成两性的可以溶解脂肪的混合微胶粒。随着食物中脂肪的摄入,它们与脂肪混合、乳化,为下一步在十二指肠吸收做好准备。胆汁酸通过肠肝再循环在体内保持稳定的水平。在整个肠道内游离和结合胆汁酸通过被动扩散的方式被吸收,在回肠末端则以主动转运形式吸收。被重吸收的胆汁酸进入血液循环系统被肝实细胞摄入重交联,再次释放进入胆汁[1],如图1。
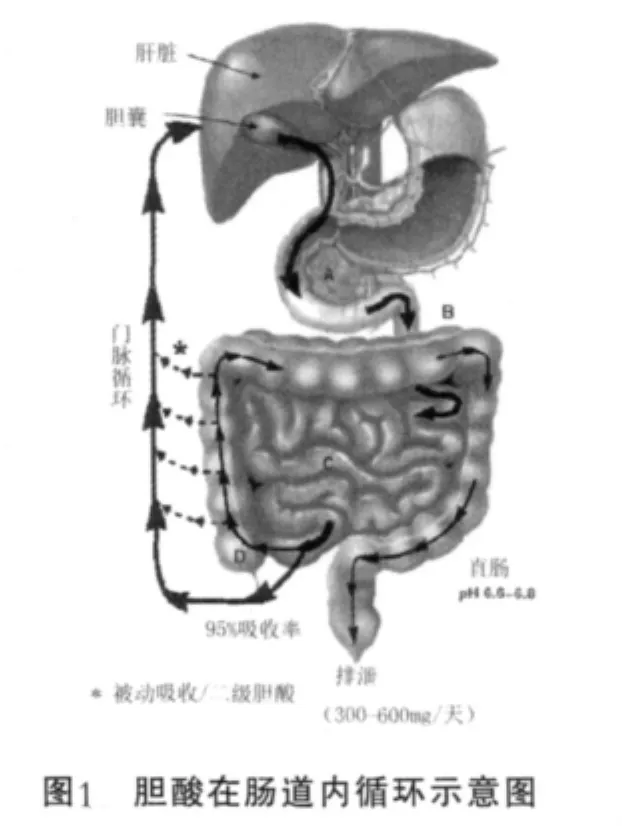
近日来很多研究开始尝试,利用哺乳动物肠道微生物群与胆盐交联,来降低高胆固醇血症患者血清中的胆固醇,以及预防正常胆固醇水平人群中高胆固醇血症的发生。肠道微生物对胆固醇调控很重要一部分是通过胆盐水解酶(Bile Salt Hydrolase,BSH)对交联胆盐水解,从而降低血清胆固醇含量来达到的。BSH催化牛磺酸或甘氨酸与甾醇基础的胆汁酸之间的氨基骨架水解。去交联的胆酸具有较低的水溶性,很容易被分泌到粪便中。由于粪便胆酸流失以及胆盐的降低使得肝脏增加胆固醇转化为胆盐来维持胆盐代谢平衡。已有实验证明,口服含胆盐水解酶的乳酸菌可以在动物及人体内降低血清中胆固醇水平。
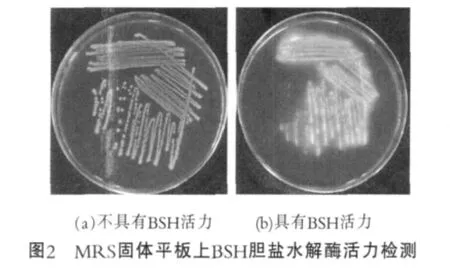
对于水解胆盐活力最常用的检测方式即在琼脂平板上观察菌落周围水解圈的出现。由于乳酸菌发酵在肠道内有时会形成局部pH较低的微环境,未交联的胆酸相比交联状态的胆酸会对细菌产生抑制作用,从而产生沉淀。这是我们在琼脂平板检测BSH活性的基础,如图2[2]。
2 胆盐水解酶bsh编码基因背景研究
BSH活性已经在乳酸菌Lactobacillus、双歧杆菌Bifidobacterium、肠球菌Enterococcus梭菌Clostridium以及 类菌体Bacteroides等细菌中被发现[3-7]。其中乳酸菌及双歧杆菌被常规认作益生菌,而类菌体梭菌以及肠球菌则是肠道共生菌。目前为止,尚未在生活环境缺乏胆盐的细菌中发现具有BSH活性的菌株。除了两株类菌体以外,其他BSH活性阳性细菌均为革兰氏阳性菌。在其他所有肠道革兰氏阴性菌中,包括大肠杆菌Escherichia coli以及肠道沙门氏菌Salmonella enterica等均未发现BSH活性,也没有找到bsh同源基因。在公共数据库中包括美国国立卫生院基因组数据库http://www.ncbi.nlm.nih.gov/以及美国Joint基因组研究所微生物基因组网站http://genome.jgi-psf.org/,我们可以检索到在已发现的益生菌中存在的编码BSH酶的基因组序列,如图3[8]。包括产气荚膜杆菌C.perfringens,植物乳杆菌La.plantarum,约氏乳杆菌La.johnsonii,长双歧杆菌Bi.Longum,两歧双歧杆菌Bi.Bifidum,青春双歧杆菌Bi.Adolescentis,以及单增李斯特氏菌Listeria monocytogenes等等.通过微生物基因组学的分析很多bsh同源基因以及预测基因也已被发现。有趣的是,在有些菌株如植物乳杆菌WCFS1中我们发现不止一个同源基因的存在[9]。同时存在多个bsh同源基因对于研究bsh基因功能具有重要意义。

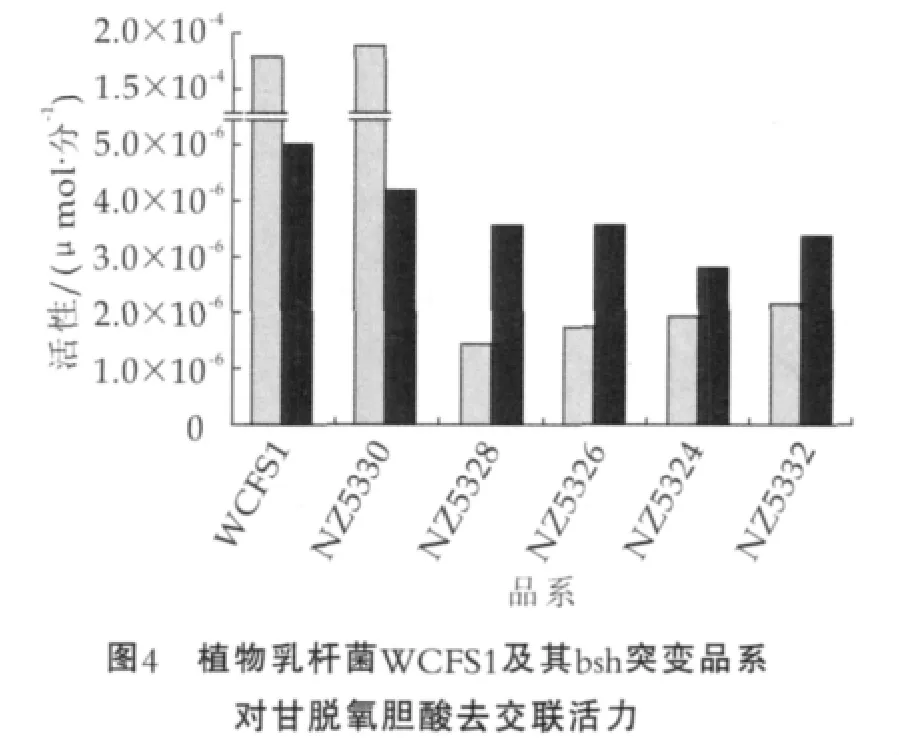
在植物乳杆菌中预测存在4个相关基因,分别命名为bsh1到bsh4。他们分散在整个基因组上,之间的氨基酸同源度从21%到39%。Lambert及Bongers等人对植物乳杆菌WCFS1中的4个bsh基因进行了深入研究,在bsh缺乏的乳酸乳球菌中分别过表达这4个bsh基因,并且构建了1个、2个、3个以及全部bsh基因敲除模型,对每个基因水解底物能力及对底物耐受性进行了相关评价。并且对若干株植物乳杆菌进行全基因组杂交(complete genome hybridization,CGH)实验结果表明,bsh2、bsh3及bsh4在植物乳杆菌各菌株间十分保守,具有重要的生理学意义,而bsh1基因则和植物乳杆菌BSH活性密切相关,如图4[9]。
3 BSH胆盐水解酶氨基酸结构及底物识别分析
在各个种属之间bsh编码区的调节和组织都有所差异。BSH酶的亚基组成、大小、最适pH值、动力学性质、底物特异性等等也变化较大。然而这些BSH却拥有一些保守的活性氨基酸位点:cysteine 2(Cys2),arginine 18(Arg18),aspartic acid 21(Asp21),asparagines 175(Asn175),and arginine 228(Arg228)(图5)。

胆盐水解酶属于胆酰甘氨酸水解酶家族(choloylglycine hydrolase family)。 青 霉 素 酰 胺 酶(penicillin amidases)也属于该家族成员,该酶可水解青霉素(penicillin)变为6-氨基青霉烷酸(6-aminopenicillinic acid),这一产物被广泛应用于半合成抗生素的工业生产中。胆盐水解酶和青霉素酰胺酶都被归类为亲核水解酶(N-terminal nucleophilic(Ntn)hydrolases),在N端带有半胱氨酸(cysteine 2,Cys2)残基。在蛋白自身酶解过程中初始的甲硫氨酸Met被去除后Cys2便成为催化中心,这也是Ntn水解酶超家族的共同点。近日产气荚膜梭菌Clostridium perfringens中的BSH酶被结晶出来,同时发现以脱辅酶(apoenzyme)及与牛磺脱氧 胆 酸 盐 (taurodeoxycholate)(TDCA;hydrolyzed product)结合成复合物形式存在。结构分析表明Cys2残基处于被亲核攻击的位置。已有实验表明Cys2的巯基(SH)基团对胆盐水解酶催化能力十分重要,在两歧双歧杆菌中将BSH带有硫醇基的Cys2置换为带羟基的亲核氨基酸Ser或Thr,则BSH失去活性[10]。在长双歧杆菌中若把BSH的Cys2置换为Ala,同样得到的是失活蛋白[11]。
图4中,横坐标从左到右依次代表:WCFS1为 野 生 型 ;NZ5330为 Δbsh2,Δbsh3,Δbsh4;NZ5328为Δbsh1,Δbsh3,Δbsh4;NZ5326为Δbsh1,Δbsh2,Δbsh4;NZ5324为Δbsh1,Δbsh2,Δbsh3;NZ5332为Δbsh1,Δbsh2,Δbsh3,Δbsh4。
同时和球形芽孢杆菌(Bacillus sphaericus)的青霉素V酰化酶(penicillin V amidase)具有高度同源性。青霉素V酰化酶的tyrosine 82(Tyr82)与BSH酶氨基酸Asn82的差异由于不同底物空间需要的差异引起。
BSH对底物胆酸的识别很可能既通过对胆酸甾类化合物的识别,也通过氨基酸集团的识别。Moser和Savage等人曾经报道在布氏乳杆菌L.buchneri JCM1069可以表达牛磺酸脱氧胆酸水解酶活性,但是却不具有牛磺酸胆酸水解酶活性[12]。牛磺酸脱氧胆酸与牛磺酸胆酸都具有牛磺酸的氨基酸部分,然而他们的甾醇部分却不同。另外在嗜酸乳杆菌NCFM中,当使bshA基因失活,该菌株水解含鹅脱氧胆酸甾醇部分 (如TCDCA和GCDCA)的胆盐活力降低[13]。不过在大多数动力学研究报告中,底物识别仍然是以氨基酸部分优先识别。大多数BSH酶水解甘氨酸交联的胆盐能力要强于牛磺酸交联的胆盐。序列分析表明虽然BSH活性位点的氨基酸十分保守,然而底物识别的位点却不是十分保守。
4 BSH与肠道的关系
胆盐水解与耐胆盐能力:因为具有去交联胆盐的能力,胆盐水解酶被认为可能具有解除胆盐毒性的作用。有4个独立研究小组利用野生型和bsh突变型细菌对胆盐水解与胆盐耐受力之间的关系进行了研究。在用亚硝基胍对食淀粉乳杆菌(Lactobacillus amylovorus)进行突变筛选中,得到一株BSH活力显著下降的突变株,在胆盐存在的情况下生长速度明显减缓[14]。在植物乳杆菌和单增李斯特菌中bsh基因产生突变,其对胆汁和胆盐十分敏感,耐受力明显下降[15]。然而BSH酶在胆汁耐受过程中的作用机理目前仍不明确。有假说提出,质子化未解离的胆盐可以如同有机酸一般通过细胞内酸化产生毒性,而具BSH活力细胞可以形成弱结合对抗物保护自己,对捕获以及转运质子造成的PH下降具有缓冲作用。
在人体内甘氨酸结合胆盐与牛磺酸结合胆盐比例为3∶1。体外实验表明,牛磺酸交联胆盐通常在任何pH情况下对细菌都仅有轻微影响,而甘氨酸交联的胆盐在酸性条件下以及bsh突变时则具有非常强的毒性。因此我们认为BSH在低pH条件下对解除甘氨酸交联胆盐毒性十分重要。当胆汁进入十二指肠时胃部会产生反酸,或者在肠道局部会产生pH较低的微环境,这时BSH意义重大。有文献报道BSHs通常会优先水解甘氨酸结合的胆盐,也有研究小组证实BSHs会倾向于微酸的环境(5到6之间)。这些报道是对这一结论有力的支持。
细菌膜特性改变:溶菌酶溶酶体(bacteriolytic enzymes lysozyme)、磷脂酶以及抗菌肽等都是高等生物肠道内体内免疫的重要执行者。细菌膜结构的组成、渗透性、流动性、净电荷以及疏水性都与细菌对寄主防御能力息息相关。有研究提出BSH可以促进胆固醇或胆汁参与细菌膜结构构成,使膜抗张强度增加,改变膜流动性及电荷等。BSH对细胞表面的改变可以增强细菌表面对体内免疫系统的抵抗能力,这种抵抗机制对建立长期感染具有重要作用。在商业中对于筛选强力BSH酶对抗BSH阴性病原菌效果显著。
促进高等动物肠道内细菌的定植:在BSH作用下胆盐去交联所产生的氨基酸,可以当作碳源、氮源以及能量来源被广泛利用。甘氨酸的代谢产物为氨和二氧化碳,而牛磺酸则可被代谢为氨、二氧化碳及硫酸盐。因此具有水解能力的菌株可能在生长时获得营养优势。Huijghebaert等人和Van Eldere等人[16]曾经观察到一株BSH阳性梭菌利用水解释放的牛磺酸作为电子受体,因此在牛磺酸和牛磺酸交联胆盐存在的情况下生长速度明显加快。在长双歧杆菌中也观察到,bsh基因总是同时伴随编码谷氨酰胺合成腺苷转移酶glnE同源基因共转录,该基因同时也是氮元素调节级联反应的一部分。
降低血清胆固醇:高胆固醇血症近年来被认为是冠状动脉心脏病的罪魁祸首。虽然已有不少治疗药物如他汀类药物(tatins)及胆酸结合剂等,但存在价格昂贵、副作用大等问题。口服益生菌目前被广泛报道具有明显降低胆固醇的能力,病例中约有22%到33%的患者胆固醇含量明显下降[17]。而小鼠实验也表明在肥胖小鼠饮食中添加益生菌可阻止胆固醇水平继续提高。益生菌降低胆固醇水平有多种途径,包括细菌对胆固醇同化作用,细菌胞壁结合胆固醇,以及短链脂肪酸发酵终产物作用等等,但BSH降低胆固醇是非常重要的途径。去交联的胆酸比交联状态胆酸更不易吸收,因此大量自由胆酸通过粪便排泄出体外。同样自由胆酸也使肠道对脂肪的溶解及吸收效率降低。因此去交联胆酸可以导致血清胆固醇含量降低,一方面为补偿通过粪便排泄损失的自由胆酸而要求胆固醇从头合成胆酸,另一方面降低胆固醇溶解度减少了肠道对其的吸收。
5 展 望
综上所述,胆盐水解对于脊椎动物脂肪代谢起着重要作用。在食物脂肪被摄入后,随着带有胆盐水解酶活力的肠道益生菌作用下,血清胆固醇被降低,肠道菌定殖能力提高,从而更好地维持肠道生态平衡。不过在众多的研究中仍存在一些不同看法。有研究者提出,由于去交联胆酸相对乳化食物脂肪和形成微粒能力降低会使得一部分正常脂肪吸收被影响。研究表明在鸡中BSH活性和鸡的发育不良有关[18],但在小鼠中未见报道。另外也有称去交联胆酸产生的二级胆酸可能对肠道功能造成一定紊乱。有一些实验也报道在胆盐耐受力与BSH活性之间未找到直接证据,但他们有的使用了牛磺酸结合胆酸而非BSH倾向结合的甘氨酸胆酸来检测BSH活性,有的则采用非等位基因菌株来研究,这对于肠道内复杂的影响因素来说不是合适的研究对象。因此这些研究并不能说明BSH真正的活性与耐胆盐能力关系。随着对BSH酶研究的越来越多,我们对于其作用机制和原理也会了解更多,这使我们阐明肠道微生物选择生存机理成为可能,同时也为发展更强健的益生菌提供了很好的研究基础。
[1]JASON M R,DAE-JOONG K,PHILLIP B H.Bile Salt Biotransformations by Human Intestinal Bacteria[J].Journal of Lipid Research,2006,47:241-259.
[2]OH H K JI Y L,SOO J L,et al.Molecular Cloning and Characterization of a Bile Salt Hydrolase from Lactobacillus acidophilus PF01[J].J Microbiol Biotechnol,2008,18(3):449–456.
[3]BATEUP J M,MCCONNELL M A,JENKINSON H F,et al.Comparison of Lactobacillus Strains with Respect to Bile Salt Hydrolase Activity,Colonization of the Gastrointestinal Tract,and Growth Rate of The Murine Host[J].Appl Environ Microbiol,1995,61:1147–1149.
[4]GRILL J P,MAGINOT-DURR C,SCHNEIDER F,et al.Bifidobacteria and Probiotic Effects:Action of Bifidobacterium Species on Conjugated Bile Salts[J].Curr Microbiol,1995,31:23–27.
[5]FRANZ C M A P,SPECHT I.Bile Salt Hydrolase Activity of Enterococci Isolated from Food:Screeningand Quantitative Determination[J].J Food Prot,2001,64:725–729.
[6]COLEMAN J P,HUDSON O L L.Cloning and Characterization of a Conjugated Bile Acid Hydrolase Gene from Clostridium Perfringens[J].Appl Environ Microbiol,1995,61:2514–2520.
[7]STELLWAG E J,HYLEMON P B.Purification and Characterization of Bile Salt Hydrolase from Bacteroides Fragilis Subsp[J].Fragilis Biochim Biophys Acta,1976,452:165–176.
[8]MÁIRE BEGLEY,COLIN H,CORMAC G M G.Bile Salt Hydrolase Activity in Probiotics[J].Applied and Environmental Microbiology,2006,1729–1738.
[9]JOLANDA M L,ROGER S B,WILLEM M D V,et al.Functional Analysis of Four Bile Salt Hydrolase and Penicillin Acylase Family Members in Lactobacillus plantarum WCFS1[J].Applied and Environmental Microbiology,2008,p.4719–4726
[10]BOGGS J M.Lipid Intermolecular Hydrogen Bonding:Influence on Structural Organization and Membrane Function[J].Biochim Biophys Acta,1987,906:353–404.
[11]TANAKA H,HASHIBA H,KOK J,et al.Bile Salt Hydrolase of Bifidobacterium Longum:Biochemical and Genetic Characterization[J].Appl Environ Microbiol,2000,66:2502–2512.
[12]MOSER S A,SAVAGE D C.Bile Salt Hydrolase Activity and Resistance to Toxicity of Conjugated Bile Salts are Unrelated Properties in Lactobacilli[J].Appl Environ Microbiol,2001,67:3476–3480.
[13]MC AULIFFE O,CANO R J,KLAENHAMMER T R.Genetic Analysis of Two Bile Salt Hydrolase Activities in Lactobacillus Acidophilus NCFM[J].Appl Environ Microbiol,2005,71:4925–4929.
[14]GRILL J P,CAYUELA C,ANTOINE J M,et al.Isolation And Characterization of A Lactobacillus Amylovorus Mutant Depleted in Conjugated Bile Salt Hydrolase Activity:Relation Between Activity and Bile Salt Resistance[J].J Appl Microbiol,2000,89:553–563.
[15]DUSSURGET O,CABANES D,DEHOUX P,et al.Listeria Monocytogenes Bile Salt Hydrolase is a Prfa-Regulated Virulence Factor Involved in The Intestinal and Hepatic Phases of Listeriosis[J].Mol Microbiol,2002,45:1095–1106
[16]HUIJGHEBAERT S M,MERTENS J A,EYSSEN H J.Isolation of a Bile Salt Sulfatase-Producing Clostridium Strain from Rat Intestinal Microflora[J].Appl Environ Microbiol,1982,43:185–192.
[17]PEREIRA D I,GIBSON G R.Effects of Consumption of Probiotics and Prebiotics on Serum Lipid Levels in Humans[J].Crit Rev Biochem Mol Biol[J].2002,37:259–281.
[18]FEIGHNER S D,DASHKEVICZ M P.Effect of Dietary Carbohydrates on Bacterial Cholyltaurine Hydrolase in Poultry Intestinal Homogenates[J].Appl.Environ.Microbiol,1988,54:337–342.
Research progress on mechanism of bile salt hydrolase in probiotics
REN Jing,WU Zheng-jun,WANG Yin-yu
(Basic Research Department,Technical Center of Bright Dairy&Food Co.Ltd.,Shanghai 200436,China)
Bile salt hydrolysis plays an important role in digestion of lipids and fat metabolism in vertebrates.We analyse the research progress of bile salt hydrolase base on the works of coding sequences,structure of amino acids and relationship between bile salt hydrolase and intestine.It is shown that the intestinal microbiota can resistant the bile salt stress and increase the ability of resistance of innate immune system.This give a great hint that we can imagine the day hypercholesterolemia be cured and serum lipids are reduced by intestinal microbiota colonization.
bile salt;bile salt hydrolase;bsh probiotics;hypercholesterolemia
Q556
B
1001-2230(2010)02-0045-04
2009-09-11
任婧(1980-),女,博士,研究方向为微生物分子生物学及遗传学。
猜你喜欢
中国食品学报(2022年12期)2023-01-12
器官移植(2022年4期)2022-07-11
电脑报(2022年25期)2022-07-05
生物信息学(2022年1期)2022-04-01
饲料工业(2021年6期)2021-04-16
食品与健康(2020年7期)2020-07-09
生物化工(2020年3期)2020-01-07
科技创新导报(2018年1期)2018-05-07
纺织导报(2017年4期)2017-05-05
中国当代医药(2015年23期)2015-03-01
