
Development of a Novel PmpD-N ELISA for Chlamydia psittaci Infection*
2016-08-15 03:24LIUShanShanCHUJunZHANGQiangSUNWeiZHANGTianYuanandHEChengKeyLabofAnimalEpidemiologyandZoonosisofMinistryofAgricultureCollegeofVeterinaryMedicineChinaAgriculturalUniversityBeijing0093ChinaTongrenPolytechnicCollegeTongr
LIU Shan Shan, CHU Jun, ZHANG Qiang, SUN Wei,ZHANG Tian Yuan, and HE Cheng,#. Key Lab of Animal Epidemiology and Zoonosis of Ministry of Agriculture, College of Veterinary Medicine,China Agricultural University, Beijing 0093, China; . Tongren Polytechnic College, Tongren 554300, Guizhou,China; 3. National and Local Engineering Research Centre for Separation and Purification Ethnic Chinese Veterinary Herbs, Tongren 554300, Guizhou, China
Original Article
Development of a Novel PmpD-N ELISA for Chlamydia psittaci Infection*
LIU Shan Shan1,2,3, CHU Jun1, ZHANG Qiang1, SUN Wei2,ZHANG Tian Yuan1, and HE Cheng1,#
1. Key Lab of Animal Epidemiology and Zoonosis of Ministry of Agriculture, College of Veterinary Medicine,China Agricultural University, Beijing 100193, China; 2. Tongren Polytechnic College, Tongren 554300, Guizhou,China; 3. National and Local Engineering Research Centre for Separation and Purification Ethnic Chinese Veterinary Herbs, Tongren 554300, Guizhou, China
Abstract
Objective Chlamydia psittaci is an avian respiratory pathogen and zoonotic agent. The wide prevalence of C. psittaci poses a threat to the poultry industry and its employees. However, few commercial kits are available for detecting avian antibodies excluding the in-house ELISA kit. In this study, we developed a novel ELISA kit for detecting antibodies against C. psittaci based on the N-terminal fragment of polymorphic outer membrane protein D (PmpD-N) as the coating antigen.
Methods The antigen concentrations, primary antibody, and cut-off value were determined and optimized. The ELISA, designated PmpD-N ELISA, was assessed for sensitivity, specificity, and concordance using sera samples from 48 experimentally infected and 168 uninfected SPF chickens.
Results The sensitivity and specificity of PmpD-N ELISA were 97.9%, 100%, respectively, while the concordance was 98.1% as compared to that of MOMP-ELISA. No cross-reaction with positive sera for other avian pathogens was found. Using PmpD-N ELISA, 799/836 clinical samples were positive, including 93.0% and 98.1% positivity in layers and broilers, respectively.
Conclusion These data indicate that indirect ELISA with PmpD-N as the antigen candidate is a promising approach for the surveillance of C. psittaci infection.
Chlamydia psittaci; Indirect ELISA; N-terminal fragment of polymorphic membrane protein D; Seroprevalence
www.besjournal.com (full text) CN: 11-2816/Q Copyright ©2016 by China CDC
INTRODUCTION
The obligate intracellular gram-negative bacterium Chlamydia psittaci (C. psittaci)causes systemic disease in psittacine birds,domestic poultry, and wild fowl[1]. In poultry, the pathogen often induces a disease called ornithosis, generally referred to as chlamydiosis in other bird species. Importantly, C. psittaci is also a zoonotic pathogen that can cause ocular adnexal lymphomas[2], pneumonia, encephalitis, endocarditis,and even death in humans[3]. Thus, chlamydiosis is associated with severe economic losses in the poultry industry, and it also poses a serious healthhazard to humans who are in close contact with infected birds[4-5]. C. psittaci strains in poultry have been reported in many countries, including Austria[6],France[7], China[8], and Germany[9]. Moreover, a high seroprevalence was recorded in Belgian and Northern French chicken farms using an indirect ELISA, with seropositivity rates of 96% and 90% in broiler and layer flocks, respectively[10]. In a recent study, C. psittaci-specific serum antibodies and C. psittaci antigens were detected in 10.0% and 26.7% of birds, respectively, suggesting that C. psittaci prevalence in Beijing is similar to that in European cities[11].
The complement fixation test (CFT) is currently used in routine diagnosis[12]. An indirect hemagglutination assay (IHA) is commercially available for detecting Chlamydia antibodies in China[13]. Unfortunately, the IHA lacks species specificity and sensitivity because of the use of inactivated whole Chlamydia elementary bodies as the diagnostic antigens. Additionally, both CFT and IHA require experienced technicians to interpret the results. The true seroprevalence of C. psittaci and potential risks are underestimated. An in-house ELISA kit based on recombinant MOMP has been developed and evaluated, with 100% sensitivity and specificity for chicken C. psittaci survey[14]. In addition to major outer membrane protein (MOMP),polymorphic membrane proteins (Pmps) are classified as autotransporters and are highly immunogenic[15]. Among the Pmps, the N-terminal fragment of PmpD (PmpD-N) is an attractive candidate because it is an immunogenic and a conserved protein among C. psittaci strains. More importantly, PmpD expression can be detected within 24 h postinfection[16]. Therefore, a reliable,sensitive, and specific test is urgently required for the preliminary survey of avian C. psittaci.
In the present study, we evaluated the sensitivity and specificity of PmpD-N ELISA for detecting avian C. psittaci antibodies. Moreover, 836 bird blood samples from 14 intensive-raising farms in seven provinces were collected to determine the seroprevalence using PmpD-N ELISA.
MATERIAL AND METHODS
Cell Culture and C. psittaci Preparation
Buffalo Green Monkey Kidney (BGMK) cells used for the propagation of C. psittaci stocks were donated by Professor WANG Cheng Ming, Yangzhou University, China. The mild-virulence C. psittaci strain CB7 (genotype A) originally isolated from a wild bird in Wuhan, China[17]was purchased from the China Institute of Veterinary Drug Control (IVDC,Beijing, China), inoculated into BGMK monolayers,and titrated according to standard protocols[18]. These isolates were inoculated into BGMK monolayers and titrated according to standard protocols. The standardized aliquots were frozen at -80 °C until use.
Serum Samples
A total of 168 SPF chickens (Vital Merial Experimental Animal Co., Ltd, Beijing, China) were selected for the immunization and comparative tests. Negativity for C. psittaci-specific antibodies was confirmed by both the in-house ELISA kit[14]and IHA kit (Lanzhou Veterinary Research Institute, Chinese Academy of Agriculture Science). Twenty-four SPF chickens were kept in an isolated facility and infected intra-tracheally with 0.1 mL of 5 × 108.5IFUs of the CB7 strain[17]. Sera from the chickens were sampled on days 0, 3, 7, and 13 post-infection.
A total of 836 chicken sera were collected randomly from 14 chicken farms in seven provinces across China, including 415 sera from laying hens aged 130-500 days and 421 sera from broiler breeder flocks aged 100-450 days. In China, 90% of all farm-raised chickens are raised in an all-in all-out production system. To ensure representative sampling, the 14 chicken farms were selected among 150 intensive poultry cooperatives listed by the Ministry of Agriculture of China using a two-stage cluster procedure. Blood samples were stored for 1 h at 37 °C. Sera were collected after centrifugation (325 ×g, 10 min, 4 °C) and stored at -20 °C before analysis.
Gene Cloning, Expression, and Purification of PmpD-N
Primers specific for pmpD-N were designed using Oligo 7 (Molecular Biology Insights, USA) based on the 6BC strain. The forward primer (PmpDF)sequence was 5'-CCGGAATTC ATGGGATCCAATGT GTTGATTTCTGGAA-3' (EcoRI site is underlined), and the reverse primer (PmpDR) sequence was 5'-CCGCTCGAGTCAAACAGCCCCACCTGTAGGAGCA-3' (XhoI site is underlined). The genomic DNA of the CB7 strain was extracted using a QIAamp@DNA Mini Kit (Qiagen Ltd., Crawley, UK), according to the manufacturer's instructions. pmpD-N was amplifiedfrom DNA by PCR using PmpDF and PmpDR. PCR amplification of pmpD-N was performed in a thermal cycler with an initial denaturation at 95 °C for 5 min,followed by 30 cycles of 94 °C for 30 s, 55 °C for 30 s,and 72 °C for 1 min, and a final extension at 72 °C for 10 min. The PCR product was cloned into the EcoRI and XhoI sites of pET30a (Novagen, Madison, WI,USA). The size and orientation were confirmed by restriction endonuclease digestion analysis and DNA sequencing of purified plasmid DNA. Then, the plasmid was transformed into Escherichia coli BL21 (DE3) (Invitrogen, CA, USA), and recombinant bacteria were selected on LB broth agar containing 50 µg/mL kanamycin. For large-scale PmpD-N production, the positive clone was grown overnight at 37 °C in 5 mL of LB supplemented with 50 µg/mL kanamycin. After overnight shaking at 37 °C, 2 mL of bacteria was added to 200 mL prewarmed LB with 50 µg/mL kanamycin. After 2 h of shaking at 37 °C, IPTG was added at a final concentration of 1 mmol/L. Following an additional incubation for 5 h at 30 °C,the bacterial cells were harvested by centrifugation. The cells were re-suspended in PBS at 2 mL/g wet weight. Lysozyme (1 mg) and benzonase nuclease (3 U/mL) were added. After incubation on ice for 30 min, the cellular debris was pelleted by centrifugation at 5000 ×g for 30 min at 4 °C. The supernatant was passed through a Ni-NTA column equilibrated with binding buffer (PBS containing 10 mmol/L imidazole). The column was then washed with washing buffer (PBS containing 20 mmol/L imidazole). The expressed PmpD-N protein was eluted three times using elution buffer (PBS containing 250 mmol/L imidazole). The purified protein was quantified using the NANODROP 2000 Spectrophotometer (Gene Ltd., USA).
Sodium Dodecyl Sulfate-polyacrylamide Gel Electrophoresis and Western Blotting Assay
Protein extracts from BL21 cells were mixed with an equal volume of reduced Laemmli sample buffer, boiled for 10 min, and separated by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The separated protein was either stained by standard methods using Coomassie brilliant blue or transferred onto polyvinylidene difluoride membranes. The proteins were then reacted with a 1:1000 dilution of the C. psittaci strain 6BC-specific polyclonal antibodies (unpublished) and a 1:4000 dilution of horseradish peroxidase (HRP)-labeled goat-anti chicken IgG(Sigma-Aldrich, Shanghai, China) for 1 h at room temperature. The membrane was developed with the chromogenic substrate 3,3′-diaminobenzidine containing 0.03% hydrogen peroxide (eBioscience,Laizee Biotech Co., Ltd, China).
Optimization of PmpD-N ELISA
The optimal concentration of the coating antigen PmpD-N and the optimal dilution of test sera were determined via checkerboard titration on a 96-well ELISA plate. Antigens were diluted from 1 to 10 µg/mL, whereas 6BC-specific polyclonal antibodies and negative sera from SPF chickens were prepared at dilutions of 1:25 to 1:800. The highest dilution of antigen was considered optimal when the optical density (OD450/630) was approximately 1.0 and the dilution provided maximum contrast between the aforementioned positive and negative sera[19-20].
The indirect ELISA was performed using the following procedure[21-22]. First, 96-well ELISA plates were coated with 100 µL/well of the purified PmpD-N protein diluted in 0.05 mol/L bicarbonate/carbonate buffer (pH 9.6) at 4 °C overnight. Subsequently, the plates were washed four times with PBS containing 0.05% Tween 20 to remove the unbound antigen, and then the wells were blocked at 37 °C for 2 h using 200 µL of blocking buffer (PBS containing 5% skimmed milk),followed by an additional washing step. Then, 100 µL of sera diluted in PBS were added into each well and incubated for 1 h at 37 °C. HRP-conjugated goat anti-chicken (HRP-IgG) antibodies (Sigma-Aldrich,Shanghai, China) were diluted 1:10,000 in PBS, and 100 µL was added to each well. After incubation and washing steps were performed as described previously, the colorimetric reaction was started by the addition of 100 µL/well 3,3,5,5′-tetramethylbenzidine (Qiagen, Stuttgart,Germany) at 37 °C for 10 min, and 2 mol/L H2SO4was used to stop color development. The plates were read using a universal Microplate Reader (Thermo Life Sciences, Shanghai, China) at 450 nm/630 nm. The cut-off value was calculated from the result of 144 sera from SPF chickens proved negative by both IHA and MOMP ELISA using the following formula mean OD + [3 × standard deviation (3SD)]. A serum sample was considered positive when its OD exceeded the cut-off value.
Evaluation of Sensitivity, Specificity, and Concordance
To evaluate the sensitivity, specificity, and concordance, serum samples infected with the CB7strain for 7 and 13 days and 168 SPF chicken sera described previously were tested using PmpD-N ELISA and in-house MOMP ELISA kits (Dr. Daisy Vanrompay, Department of Molecular Biotechnology,Faculty of Bioscience Engineering, Ghent University). The sensitivity, specificity, and concordance were calculated using the following formulae: sensitivity = [number of true positives/(number of true positives + number of false negatives)] × 100%; specificity = [number of true negatives/(number of true negatives + number of false positives)] × 100%;concordance = (the sum of positive-positive values and negative-negative values)/the total number of serum samples × 100%.
To determine whether this test has cross-reaction with other chlamydiae antibodies,PmpD-N proteins of other different chlamydiae,including Chlamydia abortus, Chlamydia felis,Chlamydia caviae, Chlamydia avium, Chlamydia gallinacean, Chlamydia pecorum, Chlamydia muridarum, Chlamydia trachomatis, and Chlamydia suis, were used to BLAST with that of C. psittaci.
Sera positive for infectious bursal disease virus (IBDV), herpesvirus of turkeys (HVT), H9N2 influenza virus (H9N2), avian metapneumovirus (aMPV), avian leukosis virus (ALV), Ornithobacterium rhinotracheale (ORT), and Haemophilus paragallinarum (Hp) were purchased from IVDC. The specific test with the aforementioned eight positive sera was further used to determine the specificity of the PmpD-N ELISA assay.
Early Infection Establishment
Chicken serum samples infected with the CB7 strain for 3 and 7 days were tested using both PmpD-N ELISA and in-house MOMP ELISA kits to test whether PmpD-N ELISA could detect early infection by C. psittaci.
RESULTS
Cloning, Expression, and Identification of PmpD-N
The amplified PCR products of the gene that encodes PmpD-N were sequenced, and the expected size of 1021 bp was observed. The PCR products were cloned into the PET-30a vector. The recombinant plasmids of the correct size were identified by PCR and gene sequencing. The pET30-PmpD-N plasmid was overexpressed successfully in E. coli host cells. Protein bands were subsequently visualized by Coomassie brilliant blue staining (Bio-Rad, Life Science, Beijing, China). SDS-PAGE revealed the PmpD-N fusion protein with an approximate molecular mass of 45 kD (including a 5.4 kD His tag) (Figure 1A), which was consistent with the expected size of the PmpD-N fusion protein. The expressed protein was then analyzed by an immunoblotting assay using 6BC polyclonal antibodies. C. psittaci-specific sera reacted with the PmpD-N fusion protein, and an approximate molecular mass of 45 kD was observed (Figure 1B). No Chlamydia-specific proteins were detected in lysates derived from pET30a-transformed E. coli cells.
Optimization of PmpD-N ELISA
Checkerboard titration was used to determine the optimal dilutions of antigens and antibodies. The optimal PmpD-N protein concentration was 4 µg/mL (Figure 2) for the coating antigen, and the dilution of the tested sera was determined to be 1:50 (Figure 2). Under these conditions, the P/N value was maximal (49.3).
Determination of the Cut-off Value
The average OD of the 144 negative sera was 0.0306, whereas the SD was 0.0048. Therefore, the cut-off OD value was 0.045 (mean + 3SD). Thus, OD values equal to or exceeding the cut-off value were confirmed to be positive, whereas those less than 0.045 were considered negative in this assay.
Sensitivity and Specificity
Serum analysis via PmpD-N ELISA revealed that 47 of 48 Chlamydia-infected chicken sera were positive, whereas one sample was negative, resulting in a sensitivity of 97.9%. By contrast, 45 of the 48 samples (93.8%) were positive using the MOMP ELISA kit. In addition, all 168 of the negative SPF samples were confirmed to be negative by both methods, yielding a specificity of 100% (Table 1).
Assessment of Cross-reactivity
Amino acid sequence alignment of different chlamydiae illustrated that the similarity between C. psittaci and other different chlamydiae ranged 34.3%-87% (Figure 3).
All OD values were below the defined cut-off point (Table 2). No cross-reactivity was found in tests with positive sera against IBDV, HVT, H9N2, aMPV,ALV, ORT, and Hp.
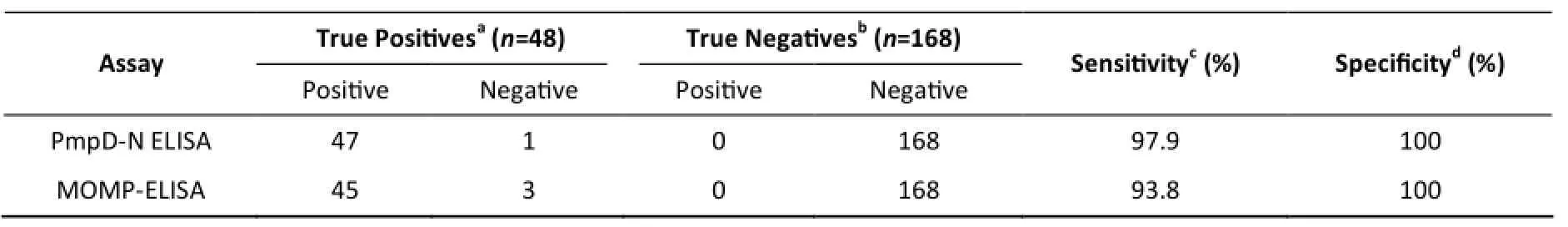
Table 1. Sensitivities and Specificities of Serological Detection of Chlamydia psittaci Antibodies in Chickens

Table 2. Specificity Test Using Positive Sera of Other Avian Pathogens
Concordance between PmpD-N ELISA and the MOMP ELISA kit
In total, 47 of the 48 positive sera identified by PmpD-N ELISA were confirmed to be positive,whereas three samples were found to be negative using the MOMP ELISA kit. On the contrary, all 168 negative sera were confirmed using the MOMP ELISA kit (Table 3). Hence, the concordance was 98.1% between the two assays.
Establishment of Early Infection
Among the 24 chicken serum samples on day 3 post-infection, 10 sera identified by PmpD-N ELISA were confirmed to be positive, whereas none was found to be positive using MOMP ELISA (Table 4). All 24 chicken serum samples on day 7 post-infection were confirmed to be positive by PmpD-N ELISA, and three sera remained negative according to MOMP ELISA (Table 4).
Clinical application of PmpD-N ELISA A total of 836 chicken serum samples from seven different provinces were tested for C. psittaci antibodies using the PmpD-N ELISA and in-house MOMP ELISA kits (summarized in Table 5). The results of the two methods were consistent. In total,799 of the 836 sera (95.6%) were determined to be positive, whereas 37 samples were found to be negative using PmpD-N ELISA. With respect to the regions, high seroprevalence was found in Northern China with the intensive raising-farm system,whereas negative samples were found in Henan,Central China. All 98 samples (100%) from Heilongjiang province, 28 samples (100%) from Liaoning province, and 212 samples (100%) from Shandong province, as well as 157 of the 159 samples (98.7%) from Tianjin, 230 of the 232 samples (99.1%) from Hebei province, and 74 of the 80 samples (92.5%) from Beijing were found to be seropositive in the study.

Table 3. Concordance between PmpD-N ELISA and MOMP ELISA for Experimentally Infected Chickens
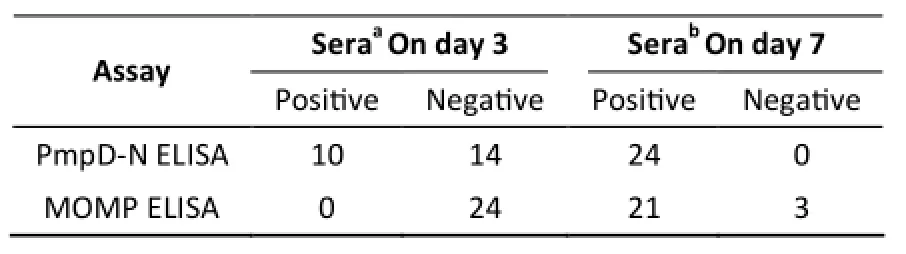
Table 4. Serological Detection of C. psittaci Antibodies in Chickens on Days 3 and 7 Post-infection
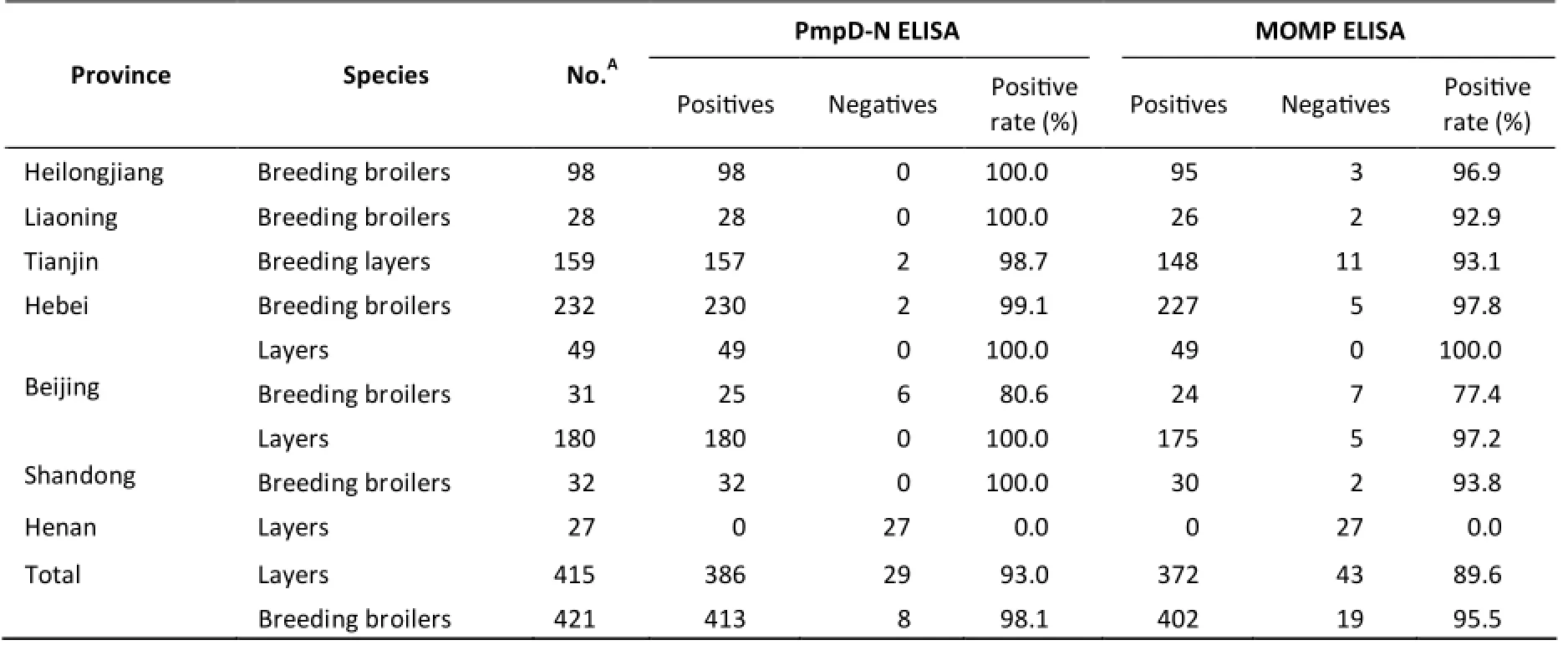
Table 5. Serological Detection of Clinical Serum Samples Using PmpD-N ELISA and MOMP ELISA
Among the chickens sampled, 386 of the 415 layers (93.0%) were found to be positive, and 413 of the 421 broiler sera (98.1%) were positive. No significant difference was observed between laying hens and breeding broilers.
DISCUSSION
The aim of this study was to develop an affordable and reliable kit for the detection of avian C. psittaci infection, as well as to develop a method to analyze the immune status of vaccinated flocks. In this study, PmpD-N ELISA was optimized as an indirect ELISA assay, which was demonstrated to be highly sensitive (97.9%) and specific (100%) for detecting C. psittaci antibodies. The concordance was 98.1% compared with the MOMP ELISA kit. Further studies demonstrated that no cross-reaction was found for sera positive for eight other chlamydiae. Therefore, PmpD-N ELISA is a promising approach for the serological diagnosis of avian C. psittaci. Its sensitivity and specificity were comparable with those of the in-house MOMP ELISA kit.
The merit of PmpD-N as the coating antigen is that it is a highly conserved fragment in the polymorphic outer protein membranes of C. psittaci,which makes it comparable with C. psittaci-specific MOMP as a diagnostic candidate. As it is difficult for us to obtain highly pure antibodies against other chlamydiae, PmpD-N proteins of other different chlamydiae were used to blast with that of C. psittaci to assess cross-reaction. The similarity of the amino acid sequence between C. psittaci and other chlamydiae ranged 34.3%-87%, suggesting that the established PmpD-N ELISA might be specific for detecting C. psittaci antibodies. In comparison with C. trachomatis and C. pneumonia, PmpD-N of C. psittaci translocates to the surface of the bacterium, where it noncovalently binds to other components of the outer membrane. Antibodies against PmpD-N were neutralizing. Recombinant PmpD-N also stimulated monocyte activation[23]. A recombinant HVT vaccine expressing PmpD-N of C. psittaci strain CB7 (rHVT-pmpD-N) was confirmed to express PmpD-N by immunoblotting and immunofluorescence, and it may be viable as a candidate dual vaccine that provides protection against both extremely virulent MDV and C. psittaci[24]. Clustering analysis resolved PmpD proteins into three distinct clades with C. abortus Pmp18D, being most similar to those originating from C. psittaci, C. felis, and C. caviae[15]. More interestingly, PmpD ELISA might be applied to detect early C. psittaci infection because of PmpD expression within 24 h of infection[16]. In the current study, C. psittaci-specific antibodies were detected on day 3 post-infection, whereas antibody positivity was not confirmed until day 7 post-infection using the in-house MOMP ELISA kit. Furthermore, PmpD-N ELISA is superior to the IHA kit, and the later kit usually gives rise to high false-positive test results because of antigenic cross-reactivity among C. abortus, C. psittaci, and C. pecorum, as well as with some gram-negative bacteria. In a recent report,12.4% of adult pet birds and 4.85% of juvenile pet birds were found to be seropositive for C. psittaci using an IHA approach[13]. In this sense, IHA is not suitable for detecting C. psittaci prevalence because of the lack of differentiation between C. psittaci and other chlamydia species in poultry in addition to poor sensitivity.
C. psittaci infection is often neglected and misdiagnosed because of the lack of a commercially available C. psittaci-specific test kit. Infection transmission from raised turkeys and ducks to humans has been previously reported[7]. Sera from Belgian and French chicken farms were also proven to be C. psittaci-positive via an indirect ELISA based on major outer membrane protein, and the positive rate was extremely high (96% in broilers and 90% in layers)[10]. The positive rate coincided with our report, which yielded 93.0% positivity in layers and 98.1% positivity in broilers. Three main factors contribute to the high seroprevalence. First, no quarantine measures are implemented for breeding boilers and breeding layers imported to China from European and other countries. A large population of breeding broilers relies on supply from international enterprises every year, and high seropositivity has been confirmed in European poultry stocks. Second,direct contacts with infected birds may be another important cause due to the lack of biosecurity measures. The infected birds could shed C. psittaci in their feces, respiratory secretions, saliva, and feather dust, and healthy birds could then be infected by this contamination[1]. Third, live vaccines are contaminated with C. psittaci, thus contributing to the transmission of C. psittaci among avian species,as C. psittaci was found in SPF eggs with the positive rates of 33.3% and 31.7%[25].
In conclusion, PmpD-N ELISA coated with the PmpD-N protein was successfully developed, and this technique is a good candidate method for C. psittaci diagnosis. More importantly, PmpD-N ELISAis comparable with MOMP ELISA with respect to sensitivity and specificity as well as concordance. It can be applied to identify C. psittaci infection.
ACKNOWLEDGMENTS
We gratefully acknowledge the correcting submission by Dr. Gordon Leitch and statistical analysis by Dr. YAN Feng Xia at Morehouse School of Medicine, Atlanta, GA, USA. The authors declare no conflicts of interest.
Accepted: April 25, 2016
REFERENCES
1. Rodolakis A, Yousef Mohamad K. Zoonotic Potential of Chlamydophila. Vet Microbiol, 2010; 140, 382-91.
2. Ferreri AJ, Guidoboni M, Ponzoni M, et al. Evidence for an Association between Chlamydia Psittaci and Ocular Adnexal Lymphomas. J Natl Cancer Inst, 2004; 96, 586-94.
3. Zhou J, Qiu C, Cao XA, et al. Construction and Immunogenicity of Recombinant Adenovirus Expressing the Major Outer Membrane Protein (Momp) of Chlamydophila Psittaci in Chicks. Vaccine, 2007; 25, 6367-72.
4. Dickx V, Geens T, Deschuyffeleer T, et al. Chlamydophila Psittaci Zoonotic Risk Assessment in a Chicken and Turkey Slaughterhouse. J Clin Microbiol, 2010; 48, 3244-50.
5. Lagae S, Kalmar I, Laroucau K, et al. Emerging Chlamydia Psittaci Infections in Chickens and Examination of Transmission to Humans. J Med Microbiol, 2014; 63, 399-407.
6. Robertson T, Bibby S, O'Rourke D, et al. Identification of Chlamydial Species in Crocodiles and Chickens by Pcr-Hrm Curve Analysis. Vet Microbiol, 2010; 145, 373-9.
7. Laroucau K, Vorimore F, Aaziz R, et al. Isolation of a New Chlamydial Agent from Infected Domestic Poultry Coincided with Cases of Atypical Pneumonia among Slaughterhouse Workers in France. Infect Genet Evol, 2009; 9, 1240-7.
8. Zhang F, Li S, Yang J, et al. Isolation and Characterization of Chlamydophila Psittaci Isolated from Laying Hens with Cystic Oviducts. Avian Dis, 2008; 52, 74-8.
9. Gaede W, Reckling KF, Dresenkamp B, et al. Chlamydophila Psittaci Infections in Humans During an Outbreak of Psittacosis from Poultry in Germany. Zoonoses Public Health, 2008; 55,184-8.
10.Yin L, Kalmar ID, Lagae S, et al. Emerging Chlamydia Psittaci Infections in the Chicken Industry and Pathology of Chlamydia Psittaci Genotype B and D Strains in Specific Pathogen Free Chickens. Vet Microbiol, 2013; 162, 740-9.
11.Ling Y, Chen H, Chen X, et al. Epidemiology of Chlamydia Psittaci Infection in Racing Pigeons and Pigeon Fanciers in Beijing, China. Zoonoses Public Health. 2014.
12.Moore FM, McMillan MC, Petrak ML. Comparison of Culture,Peroxidase-Antiperoxidase Reaction, and Serum Latex Agglutination Methods for Diagnosis of Chlamydiosis in Pet Birds. J Am Vet Med Assoc, 1991; 199, 71-3.
13.Cong W, Huang SY, Zhang XX, et al. Chlamydia Psittaci Exposure in Pet Birds. J Med Microbiol, 2014; 63, 578-81.
14.Verminnen K, Van Loock M, Hafez HM, et al. Evaluation of a Recombinant Enzyme-Linked Immunosorbent Assay for Detecting Chlamydophila Psittaci Antibodies in Turkey Sera. Vet Res, 2006; 37, 623-32.
15.Wheelhouse NM, Sait M, Aitchison K, et al. Processing of Chlamydia Abortus Polymorphic Membrane Protein 18d During the Chlamydial Developmental Cycle. PLoS One, 2012; 7,e49190.
16.Kiselev AO, Skinner MC, Lampe, MF. Analysis of Pmpd Expression and Pmpd Post-Translational Processing During the Life Cycle of Chlamydia Trachomatis Serovars a, D, and L2. PLoS One, 2009; 4, e5191.
17.Chu J, Sun R, Wu Z, et al. Whole-Genome Sequences of Low-Virulence Strain Cb3 and Mild Strain Cb7 of Chlamydia Psittaci. Genome Announc, 2014; 2.
18.Yang J, Ling Y, Yuan J, et al. Isolation and Characterization of Peacock Chlamydophila Psittaci Infection in China. Avian Dis,2011; 55, 76-81.
19.Bakheit MA, Schnittger L, Salih DA, et al. Application of the Recombinant Theileria Annulata Surface Protein in an Indirect Elisa for the Diagnosis of Tropical Theileriosis. Parasitol Res,2004; 92, 299-302.
20.Zheng FY, Lin GZ, Qiu CQ, et al. Serological Detection of Bovine Ephemeral Fever Virus Using an Indirect Elisa Based on Antigenic Site G1 Expressed in Pichia Pastoris. Vet J, 2010; 185,211-5.
21.Kumar JS, Parida M, Lakshmana Rao PV. Development & Evaluation of Biotinylated DNA Probe for Clinical Diagnosis of Chikungunya Infection in Patients' Acute Phase Serum & Csf Samples. Indian J Med Res, 2013; 138, 117-24.
22.Arya P, Ponmariappan S, Singh L, et al. Development of Elisa Based Detection System for Lethal Toxin of Clostridium Sordellii. Indian J Med Res, 2013; 137, 1180-7.
23.Wehrl W, Brinkmanng V, Jungblut PR, et al. From the inside out--Processing of the Chlamydial Autotransporter Pmpd and Its Role in Bacterial Adhesion and Activation of Human Host Cells. Mol Microbiol, 2004; 51, 319-34.
24.Liu S, Sun W, Chu J, et al. Construction of Recombinant Hvt Expressing Pmpd, and Immunological Evaluation against Chlamydia Psittaci and Marek's Disease Virus. PLoS One, 2015;10, e0124992.
25.Chen X, Yuan JL, Zhang S, et al. Preliminary Survey of Spf Chickens and Vaccines Contaminated by C. Psittaci. China animal husbandry and veterinary medicine, 2010; 37, 4.
Biomed Environ Sci, 2016; 29(5): 315-322 10.3967/bes2016.041 ISSN: 0895-3988
*This work was supported by the National Natural Science Foundation of China under Grant No. 31272542; and Ministry of Science and Technology (MoST) grant 2012AA101302 (HE Cheng).
#Correspondence should be addressed to Professor HE Cheng, Tel/Fax: 86-10-62733613, E-mail: hecheng@cau.edu.cn
Biographical note of the first author: LIU Shan Shan, female, born in 1986, PhD, majoring in preventive veterinary medicine.
January 18, 2016;
 Biomedical and Environmental Sciences2016年5期
Biomedical and Environmental Sciences2016年5期
- Biomedical and Environmental Sciences的其它文章
- Evaluation of Six Recombinant Proteins for Serological Diagnosis of Lyme Borreliosis in China*
- Viral Etiology Relationship between Human Papillomavirus and Human Breast Cancer and Target of Gene Therapy
- Whole Genome Sequencing and Comparisons of Different Chinese Rabies Virus Lineages Including the First Complete Genome of an Arctic-like Strain in China*
- The Status and Associated Factors of Successful Aging among Older Adults Residing in Longevity Areas in China*
- Cognitive Training in Older Adults with Mild Cognitive Impairment
- Dietary Exposure to Benzyl Butyl Phthalate in China*